7. Поровые устройства грибов
Поровые аппараты грибов не могут быть отнесены к самостоятельным органеллам клетки, так как являются частью поперечной клеточной септы. Самостоятельное значение поровых устройств как лабильных клеточных органелл заключается в том, что они либо разрешают обмен клеток питательным материалом или даже органеллами вплоть до ядер, либо полностью замыкают их от соседствующих с ними сегментов мицелия. Поскольку у грибов других средств связи между клетками, таких как плазмодесмы, не существует, то она возможна для них только через поры септ, и состояние мицелия с замкнутыми пробками может рассматриваться как одноклеточное, а сам мицелий - как популяция одноклеточных форм, что может привести к далеко идущим выводам в плане экологии и генетики.
Сложные долипоры базидиальных грибов обратили на себя внимание давно, и изучение их показало, что они снабжены боковыми выростами - парентосомами, прикрывающими сверху отверстие септы двусторонними колпачками (Gull, 1978) и затыкающими туннель поры пробками, состоящими из белковых филаментов, растворяемых протеолитическими энзимами (Flegler et al., 1976). В вегетативном мицелии голобазидиальных агариковых грибов долипоры обычно закрыты пробками и колпачками парентосом, тогда как в гимении плодовых тел в обрамлении долипор наблюдаются перфорации, а пробки и колпачки отсутствуют (рис. 7.10, а, б) и обмен цитоплазмой и органеллами между сегментами гиф плодового тела протекает беспрепятственно. Закрытая колпачками и пробками зона остается только на границе гимения и субгимения, причем колпачок и неперфарированное обрамление сохраняются только на пограничной стороне субгимения (Burnett, 1968). Это правило соблюдается также у сумчатых дискомицетов, у которых изолирующие устройства пор располагаются обычно между гимением и субгимением с пограничной стороны последнего (Камалетдинова, Васильев, 1982). У дейтеромицетов такие изолирующие устройства наблюдаются в местах отчленения конидий или между отдельными клетками в многоклеточных конидиях, исчезая при их прорастании (Murrey, Maxwell, 1976). Они встречаются и в гифах мицелия продуцента пенициллина Penicillium chrysogenum (см. атлас Kurylowicz et al., 1980, рис. 42).
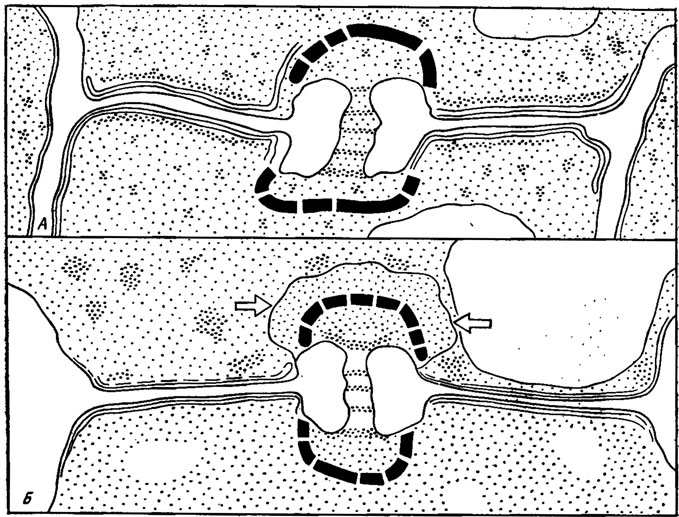
Рис. 7.10. Схема долипор плодового тела базидиомнцета Agrocybe praecox: a - септа гимениальной области; б - септа в пограничной области между гимением и субгимением. Электронно-плотный колпачок присутствует только со стороны субгимеиия (стрелки); такие же колпачки окружают с обеих сторон септы долипоры, расположенные в ткани остального субгимеиия (Gull, 1978)
У сумчатых грибов из порядка дискомицетов имеется чрезвычайное разнообразие замыкающих поровых устройств (Камалетдинова, Васильев, 1982/рис. 9). Их известно не менее пяти: диафрагмы, куполовидные тельца, тельца Воронина, полосатые тельца и пробки. Для более примитивной группы иноперкулятных дискомицетов характерно лабильное и наименее плотное замыкание поры тельцем Воронина, которое происходит, возможно, с помощью сократительных движений микрофиламентов. Тельца Воронина дисковиХной формы в такой закрытой поре могут быть расположены по нескольку штук (рис. 7.11). Такие изоляции весьма лабильны. Они легко открываются и снова закрываются в зависимости от физиологического состояния клеток гиф, регулируя при этом ток цитоплазмы или поступление питательных веществ из одного сегмента гиф в другой. Формирование изолирующих устройств в виде телец Воронина не связано со старением гиф или только с экстремальными условиями среды, как наблюдалось ранее у Neurospora (Trinci, Collinge, 1974) и у Ascodesmis sphaerospora (Brenner, Carrol, 1968). Так, в активно растущих жизнеспособных колониях Erysiphe graminis до 80% септальных пор закрыты пробками из телец Воронина (МсКееn, 1971), что говорит о их роли в специализированной дифференциации обмена.
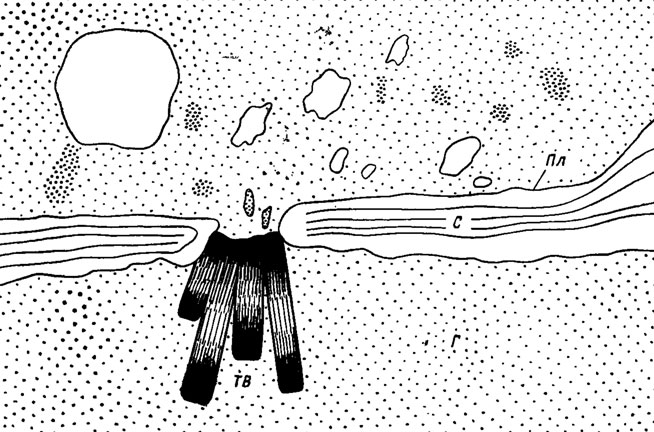
Рис. 7.11. Схема септальной поры медуллярного эксципула плодового тела дискомицета Morchella conica, закрытая пластинчатыми тельцами Воронина (ТВ): С - септа; Пл - плазмалемма (по Камалетдиновой, Васильеву, 1982)
Изолирующее устройство оперкулятных дискомицетов и иноперкулятных форм включает кроме телец Воронина еще и диафрагму - линзообразное герметичное образование в просвете поры, происходящее от пролиферации септы и ограниченное с двух сторон плазматической мембраной. Разрастание и перекрывание диафрагмы фибриллярным или гранулярным материалом формирует еще более герметичное куполовидное тельце. В ряде случаев диафрагма замещается полосатым тельцем, фильтрующим устройством, составленным из стопок дисков или цилиндров полисахаридной природы. Все эти образования могут комбинироваться до формирования полной пробки, обеспечивающей наиболее герметичную изоляцию клетки (рис. 7.12/Камалетдинова, Васильев, 1982/рис. 11, 15).
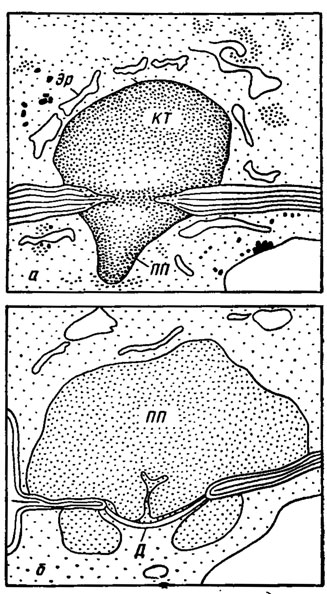
Рис. 7.12. Схема полных поровых пробок в основании сумок плодовых тел дискомицетов: а - куполовидное тельце (КТ) с полной пробкой (ПП) в поре основания сумки Scutellinia scutellata; Эр - эндоплазматический ретикулум; б - полная пробка (ПП) с диафрагмой (Д) в основании молодой сумки Peziza varia (по Камалетдиновой, Васильеву, 1982)
Изолирующие и фильтрующие устройства имеют несомненное отношение к приспособительной онтогенетической дифференциации, которая так развита в царстве грибов.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'