8. Локализация в клеточных органеллах звеньев обмена веществ
Для выяснения локализации в различных клеточных органеллах разных звеньев обмена веществ и соответствующих им ферментных систем оказался очень полезным предложенный Залокаром (Zolokar, 1965) метод нативного центрифугирования содержимого живых гиф грибов в ультрацентрифуге при центробежной силе от 10000 до 100000 г/см. Преимущество метода заключается в отсутствии артефактов, которые неизбежно бывают при работе с гомогенатами, и в возможности проверить их отсутствие с помощью контроля жизнеспособности отцентрифугированных гиф по продолжению их роста на питательной среде.
Цитохимический анализ составных частей клетки показал следующее расположение в клеточных органеллах грибов мест биосинтетических реакций клетки (Zolokar, 1965/рис. 7.13, 7.14).

Рис. 7.13. Схематическое изображение результатов цитохимического анализа распределения протеинов, иуклепротеидов, липидов и углеводов в клеточных органеллах Neurospora crassa после центрифугирования мицелия (направление центробежной силы слева направо); 1 - распределение органелл по длине гифы; ЛВ - липиды и вакуоли; С - супернатаит (цитоплазма); Я - ядра; М - митохондрии; Эр - эндоплазматический ретикулум и рибосомы; Г - гликоген; 2 - реакция Паули на протеины; 3 - реакция Сакагучи на протеины (аргинин); 4 - реакция Барнетта и Зелигмана (на SH-связи протеинов); 5 - окраска РНК (акридин - оранж); 6 - окраска ДНК ядер по Хюбшмаину (азурА - О2); 7 - реакция Бауэра на гликоген; 8 - реакция Гочкяса на углеводы (периодат - Шнфф); 9 - реакция на жиры (судан IV); 10 - окраска на липиды (тетраксид осмия); (по М. Zalolcar, 1965)
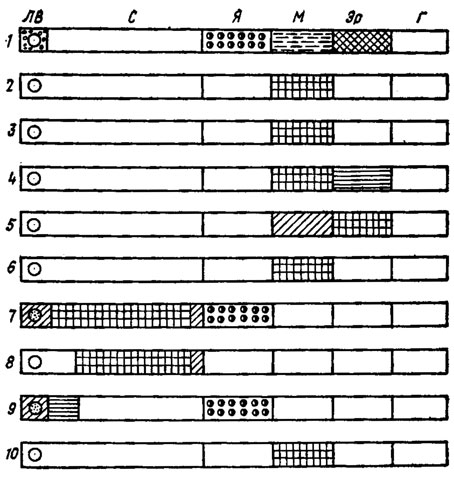
Рис. 7.14. Схематическое изображение результатов цитохимического анализа распределения ферментов в клеточных органеллах Neurospora crassa после центрифугирования мицелия (направление центробежной силы слева направо); 1 - распределение органелл по длине гифы их обозначения как на рис. 7.13); 2 - сукцииатдегидрогеназа (тетрозолий); 3 - цитохромоксидаза (НАДН); 4 - пероксидаза (бензидин); 5 - пероксидаза (десятиминутная); 6 - β-галактозндаза; 7 - щелочная фосфатаза, реакция Гомори (аденозии-3-фосфат); 8 - щелочная фосфатаза (β-нафтилфосфат, диаэо-синяя); 9 - кислая фосфатаза реакция Гомори (глицерофосфат); 10 - кислая фосфатаза (α-нафтилфосфат, диазо-синяя) (по М. Zalokar, 1966)
В ядре локализуется синтез ДНК, РНК, НАД, располагаются энзимы нуклеозидфосфорилаза, аргиназа, ДНК- и РНК-полимераза и НАД-синтетаза, локализуется синтез протеинов. Локализация синтеза РНК в ядре подтверждена с помощью сочетания центрифугирования живых гиф с ауторадиографией.
В митохондриях локализованы:
1) синтез всех кислот цикла Кребса; 2) обмен жиров и липидов и их синтез; 3) окислительное фосфорилирование, локализация фосфатаз и субстратов их действия (АМФ, АДФ, АТФ); 4) дыхательная цепь ферментов, НАД, НАДФ, флавиновые энзимы, цитохромы, цитохромоксидаза, цитохромредуктаза; 5) пероксидаза; 6) ферменты, участвующие в аминировании, в образовании пептидных связей, протеолитические ферменты, трансаминазы, система ферментов, производящих синтез оротовой кислоты и предшественников пуринов и т. д.; 7) некоторые ферменты углеводного обмена (β-галактозидаза).
Рибосомы представляют собой место синтеза белка, содержат РНК и ацилирующие энзимы, холин и витамин А-эстеразы, апиразу. В идентичной по размерам частиц микросомальной фракции содержатся ферменты переработки углеводородов и липидов, энзимы, связанные с явлением люминесценции.
В цитоплазматической гидрофильной фракции содержатся главным образом энзимы, участвующие в процессах гликолиза. К числу ферментов, локализованных в пределах клеточной мембраны, относятся такие гидролитические энзимы, как инвертаза, целлобиаза, трегалаза и мальтаза.
Ознакомление с локализацией процессов обмена в клеточных структурах имеет не только научный интерес, но является полезным и для практических отраслей микологии. Наблюдение за поведением клеточных органелл может нести контрольную службу в процессах ферментации или борьбы с патогенами, позволяя прогнозировать или подтверждать направление процессов обмена гриба в сторону либо стимуляции, либо угнетения как общего развития, так и интересующего обменного шунта. Изыскания в области стимуляции развития органелл, являющихся материальной основой обменных процессов, должны приводить к улучшению этих процессов в целом и их регулированию. Именно в этой форме: представлял себе дальнейшее развитие подобного направления: исследований их зачинатель М. Н. Мейсель (1950).
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'