Глава 6. Ферменты грибов
Питание и обмен веществ у живых организмов были бы невозможны без специфических органических катализаторов - ферментов, и в особенности это касается организмов гетеротрофов, которые используют для своего питания сложные органические соединения, находящиеся в окружающей их среде. Эти органические соединения очень часто, если не в большинстве случаев, представляют собой нерастворимые в воде полимеры, которые необходимо при их использовании расщепить на отдельные молекулы - мономеры. Для этого требуется широко развитая способность выделять в окружающую среду экзоферменты, приспособленные к находящемуся в ней субстрату. Другими словами, гетеротрофным организмам необходимо обладать способностью изменять в зависимости от условий среды качественный и количественный состав синтезируемого ими комплекса ферментов.
Ферменты не распределены равномерно по всей клетке, а в большинстве случаев связаны с определенными ее структурами. Это показали уже самые первые опыты по суперцентрифугированию клеточных гомогенатов. При этом нижний слой гомогената, содержащий митохондрии, показал в аппарате Варбурга поглощение кислорода, измеряемое 2500 условными единицами, средний слой, содержащий рибосомы - 60 единицами, а у не содержащего субклеточных структур верхнего слоя поглощение кислорода отсутствовало полностью. Такой прозрачный цитоплазматический слой обычно содержит только гидролитические ферменты и большую часть ферментов путей гликолиза, тогда как ферменты цикла Кребса, дыхательной цепи и большинство синтетаз содержатся в митохондриях, микросомах, рибосомах, мембранах и в ядрах клеток. Гидролитических ферментов может оказаться больше, чем в клетке, в окружающей среде, куда они интенсивно выделяются.
Однако чаще это происходит не сразу после их биосинтеза, сопровождающего рост продуцента. Первоначально они накапливаются в мицелии, как связанные в цитоплазматической мембране. Участвующие в их биосинтезе связки рибосом и соответствующая информационная РНК локализуются на внутренней стороне плазматической мембраны (рис. 6.1). Формируемый ими фермент локализуется внутри нее и представляет собой в это время высокомолекулярный фосфолипопротеин, содержащий одновременно липофильную и гидрофильную с терминальной NН2-группой цепи, способствующие ее транспортировке через мембрану. Окончательно сформированный энзим попадает далее в периплазматические пузырьки - мезосомы, причем его молекулярная масса падает в ~20 раз, видимо, за счет потери транспортных устройств, отщепляемых эндопептидазами. Энзим принимает при этом форму энзиматически активного глобулярного белка и транспортируется на наружную поверхность мембраны и в окружающую среду (Borris, 1981).
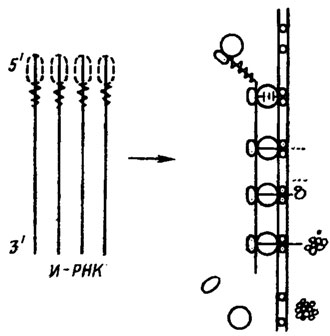
Рис. 6.1. Гипотетическая модель синтеза и секреции экзоэнзима. Сигнальный кодой информационной РНК обозначен как область зигзага, сигнальная последовательность в начале нативного протеина - как пунктирная линия. Направо - изменение конформации и выделение через клеточную мембрану в среду синтезированного энзима (Borris, 1981)
Сложный процесс синтеза и транспорта таких энзимов, естественно, разобщает во времени сопровождающий рост общий биосинтез белков и выделение и накопление гидролаз в культуральной среде. Именно с этим связано наблюдаемое у грибов резкое различие в длительности жизни информационной РНК, кодирующей биосинтез секреторных гидролаз, по сравнению с кодирующей внутриклеточные энзимы, что, вероятно, зависит от течения формирования секреторных энзимов в мембрано-связанном состоянии.
Связанное с полимерами мембран состояние способствует сохранности стабильности не только кодирующей синтез ферментов информационной РНК, но и самих энзимов, что в последнее время широко используется для их стабилизации.
Среди ферментов различают конститутивные, формирующиеся с самого начала онтогенеза, и адаптивные, появляющиеся впоследствии в зависимости от условий окружающей среды. В случае гидролитических адаптивных ферментов, расщепляющих высокомолекулярные субстраты (полисахариды, белки и т. п.), индукторами их появления могут быть либо их субстраты, либо иногда мономерные продукты производимых ими реакций. Последнее отмечалось для некоторых ферментов, гидролизующих полисахариды, например, индуктором некоторых пектиназ является галактуроновая кислота. Субстрат реакции нередко индуцирует также формирование ферментов, участвующих в детоксикации. Так, принадлежащий к эстеразам фермент трихотециназа начинает образовываться у почвенных грибов при наличии в среде субтоксических доз антибиотика трихотецина.
Даже конститутивные энзимы, к которым относится большинство ферментов основного обмена, в онтогенезе грибов формируются не сразу, а в определенной последовательности, и кроме того, набор их в зависимости от возраста культуры сильно меняется как по количественным соотношениям, так и качественно. Это можно видеть из целого ряда примеров, часть из которых уже приводилась при обсуждении путей гликолиза у грибов. Было установлено, что изменения в наборе дыхательных энзимов, включающих гексокиназу, глицеральдегид-3-фосфатдегидрогеназу, дегидрогеназы кислот цикла Кребса и ряд других, и в их активности наблюдаются уже при прорастании спор ряда плесневых грибов, как Trichoderma viride, Aspergillus niger, Penicillium atrovenatum, P. oxalicum. В процессе образования ростовых трубок активность ряда ферментов, особенно имеющих в качестве кофермента НАД, возрастала в 51-200 раз.
Аналогичная картина наблюдалась при развитии у грибов органов плодоношения. При развитии шляпки плодовых тел Schizophyllum commune в ней значительно возрастала активность R-глюканазы, расщепляющей содержащийся в оболочках его клеток полисахарид с одной-тремя связями мономеров глюкозы, R-глюкан. Особенно активизировался этот адаптивный энзим при недостатке в среде глюкозы (Wessels, 1966). Активность многих ферментов меняется не только на протяжении всего онтогенеза или его узловых этапов, но и при таких краткосрочных периодических процессах, как деление клеточных ядер. Так, у базидиомицета Clavatia cyatiformis в гифах, находящихся в состоянии мейоза (редукционного деления), резко возрастала активность фосфатазы (Blumer, Li-Yu-Ten, 1966). Сильно изменялось также содержание окислительных и других ферментов в процессе развития мицелия Penicillium chrysogenum при ферментации пенициллина (Маттисон, 1956) и концентрация амилазы в погруженной культуре Aspergillus niger. Примеры подобного рода могут быть значительно расширены.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'