3. Жирорастворимые витамины
К жирорастворимым витаминам относят витамин А и группу витамина D, а также витамины К, Е и Q. Все эти соединения относятся к продуктам обмена шунта терпенов, хотя и синтезирующихся на разных его этапах: А - на пути биосинтеза каротиноидов, D-стероидов, Q, К и Е - при частичной циклизации фарнезола или сквалена. Представители почти всех этих витаминов (кроме К и Е) обнаружены у грибов, и они синтезируют их настолько хорошо, что потребность в них у этих организмов, как правило, отсутствует.
Витамин А и каротиноиды. Витамин А был первым из изученных витаминов и представляет собой ровно половину молекулы β-каротина, расщепление которого по шестой из его одиннадцати двойных связей дает две молекулы витамина А (рис. 5.7).
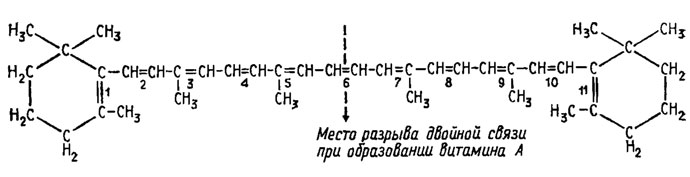
Рис. 5.7. Структура β-каротина и формирование из него витамина А
α-Каротин дает при расщеплении только одну молекулу этого витамина, а ликопин ни одной, так как для формирования структуры витамина А необходимо присутствие циклической β-иононовой группировки, которых в α-каротине одна, а в ликопине они отсутствуют. Грибы обычно очень хорошо синтезируют каротиноиды и поэтому в источниках витамина А не нуждаются. Биосинтез каротиноидов у грибов широко изучался Т. Гудвином (1954, 1962), которому удалось показать его путь через конденсацию пентенилфосфата и фарнезилфосфата при инкубации этих веществ с дрожжевым экстрактом, в результате чего был получен геранилгераниол - соединение, представляющее собой половину молекулы алифатических каротиноидов типа ликопина. Предположительно, первый из формирующихся алифатических каротиноидов - фитоин - образуется путем конденсации двух молекул геранилгераниола.
Дальнейший путь их биосинтеза протекает через ступенчатое окисление (Weete, 1980) с потерей на каждой ступени двух протонов, сопровождающейся образованием новой двойной связи (рис. 5.8). При этом на третьей ступени окисления образуется нейроспорин, а на четвертой ликопин, концевые группы цепей которых претерпевают циклизацию с образованием β-зеакаротина (из нейроспорина) или γ-каротина (из ликопина), формирующего на конечном этапе β-каротин (рис. 5.9). Все эти виды каротиноидов были обнаружены у грибов, однако необходимо отметить, что типичной для них является широкая представленность веществ с кислой природой, подобных каротиноидному пигменту раков, астаксантину. Относительно сильно окисленные соединения - ксантофиллы - у грибов сравнительно редки, но сильно восстановленных и метоксилированных каротиноидов встречается много (Гудвин, 1954, 1962). У грибов обнаружено до 7 каротиноидов с циклической структурой концевых групп и 11-12 двойными связями (рис. 5.10). Среди них особенно широко распространены β-, α- и γ-каротины и несколько реже δ-каротин (у базидиальных грибов). К той же группе относятся гидроксилированные каротиноиды типа ксантофилла, как рубиксантин, встречающийся также у базидиомицетов, метоксилированный торулин и каротиноид с деградированным кольцом и карбоксильной группой, торулародин, встречающиеся у Rhodotorula и ряда других сумчатых и базидиальных грибов.
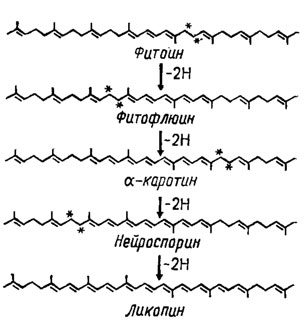
Рис. 5.8. Путь превращения фитоина в ликопин через промежуточные стадии его окисления. Звездочки указывают атомы углерода, теряющие водород на следующей стадии окисления (Weete, 1980)
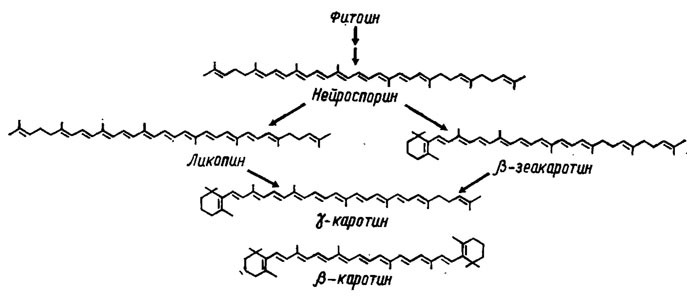
Рис. 5.9. Биосинтез β-каротина и γ-каротина у грибов из алифатических форм каротиноидов (Weete, 1980)
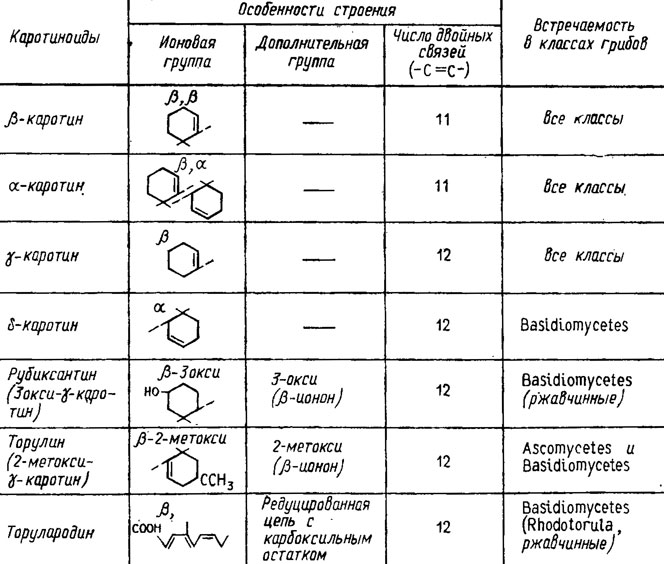
Рис. 5. 10. Каротиноиды грибов с циклической структурой (Гудвин, 1954; Weete, 1980)
У грибов также встречается много каротиноидов (порядка 8) типа ликопина с алифатической структурой (рис. 5.11), отличающихся друг от друга разной степенью восстановленности или наличием гидроксильных (ликоксантин) или метоксильных (родовиоласцин) групп. Ряд соединений из этой группы составляет последовательную прогрессию, в которой каждый последующий компонент отличается от предыдущего выпадом одной двойной связи при включении в молекулу двух атомов водорода: ликопин (13 С = С) → нейроспорин (12 С = С) → β-каротин (11 С = С) → фитофлуин (10 С = С) → фитоин (9 С = С). Фитофлуин и фитоин, в отличие от других каротиноидов с их желтой, оранжевой или красной окраской, бесцветны, но фитофлуин флуоресцирует в ультрафиолетовом свете, а фитоин поглощает его при длине волны около 220 нм. Все это зависит от потери обусловливающих окраску и длину волны отражаемого света двойных связей.
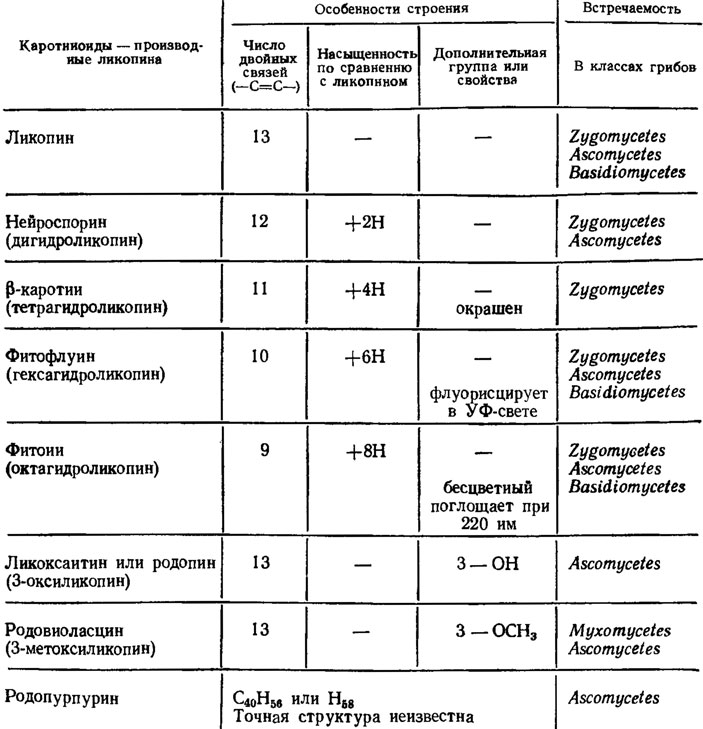
Рис. 5.11. Каротиноиды грибов - производные ликопина (Гудвин, 1954; Weete, 1980)
Количество бесцветных каротиноидов нередко в сотни раз превышает количество окрашенных, например у Rhodotorula gracilis (Вечер, Куликова, 1968). Все эти алифатические каротиноиды встречаются преимущественно у зигомицетов и аскомицетов, хотя некоторые из них были найдены и у базидиальных грибов (ликопин, фитофлуин, фитоин) и миксомицетов (родовиоласцин/Гудвин, 1954). Каротиноиды часто встречаются у грибов, хотя у некоторых они не найдены, а разнообразие их не меньшее, чем у высших растений. Они обильны во всех таксономических группах, только разнообразие их связано в большей мере с высшими грибами из сумчатых и базидиальных и в меньшей мере представлено у миксомицетов и хитридиомицетов. В определенных таксономических группах встречаются сходные каротиноиды. Например, у розовых дрожжей Rhodotorula и Sporobolomyces - β-каротин и торулин, а у всех видов Rhodotorula еще и торулородин. Всем видам рода Allomyces свойствен γ-каротин.
Образуют каротиноиды многие дейтеромицеты, а также сумчатые грибы и особенно базидиомицеты, которые пока изучены весьма слабо. Из зигомицетов наиболее обильно образуют каротиноиды виды родов Phycomyces, Pilobolus, Choanephora (Blakeslea) trispora и другие виды Choanephora (Милько, Белякова, 1969).
Поскольку каротиноиды могут служить источником чрезвычайно важного для всех организмов витамина А, большое внимание было уделено изучению условий их биосинтеза. Для образования каротиноидов обычно бывает полезен сдвиг соотношения между азотом и углеродом в среде в сторону большего преобладания углерода. Не все источники углерода одинаково хороши для этого биосинтеза. Например, для биосинтеза каротиноидов у Sporobolomyces рафиноза и мальтоза оказываются лучшими источниками углерода, чем глюкоза, а глицерин - лучшим, чем маннит. Образование каротиноидов у этого вида стимулировали янтарная кислота и некоторые аминокислоты, например, сходный с изопренами по структуре и числу атомов углерода валин, а также аспарагин, аргинин и глютаминовая кислота (Бобкова, 1965). Благоприятный эффект добавки 0,05% таких аминокислот, как лейцин, глицин, глютаминовая кислота и аспарагин для синтеза каротиноидов, был обнаружен также у Choanephora trispora (Дедюхина, Бехтерева, 1968).
При введении в среду неорганических источников азота Sporobotomyces лучше синтезировал каротин на среде с азотнокислым калием, поскольку каротиногенезу способствует слабощелочная или нейтральная реакция среды. Однако на среде с сернокислым аммонием лучше поддерживался рост мицелия, вследствие чего она оказывалась в общем более благоприятной для биосинтеза каротина (Бобкова, 1965). Биосинтез каротиноидов у Choanephora trispora лучше всего протекает при росте на комплексных органических средах, например на смеси из гидролизатов соевой и кукурузной муки (Колот, 1969; Бехтерева и др., 1969).
Кроме сходных по структуре с изопренами аминокислот, например валина, и растительных масел, например хлопкового, в качестве стимуляторов синтеза каротиноидов в среду оказалось полезным добавлять после 48 ч роста их предшественники, сходные по структуре с β-иононом или сам β-ионон в концентрациях от 0,1 до 0,5% (см. рис. 5.12).
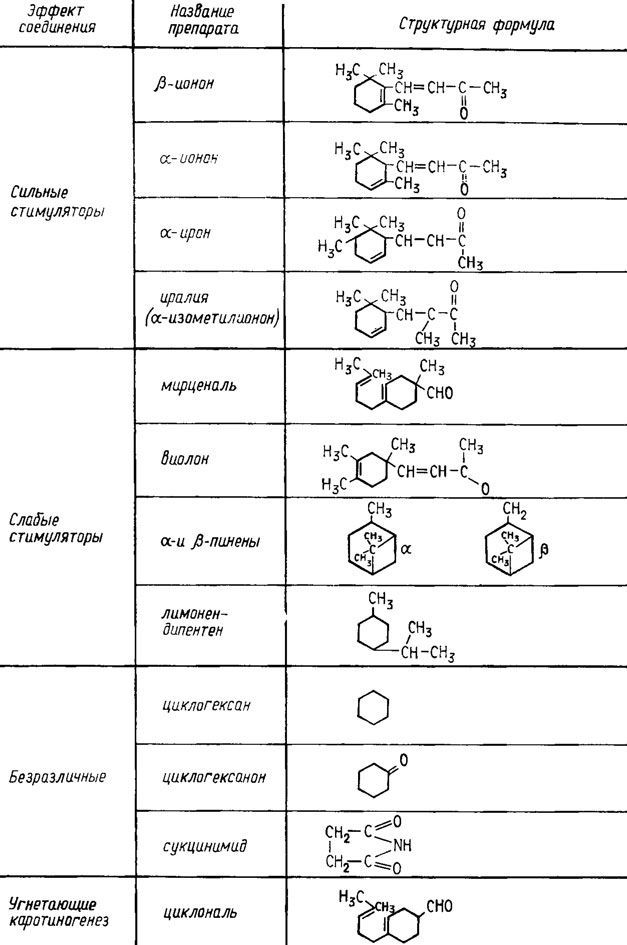
Рис. 5.12. Терпеновые соединения, влияющие на каротиногенез у Choanephora (Blakeslea) trispora (Колот и др., 1969)
Другим сильным стимулятором каротиногенеза, выделяемым из культуральной жидкости совместной культуры (+) и (-) штаммов Choanephora trispora является так называемая триспоровая кислота или фактор β (Бехтерева и др., 1969):
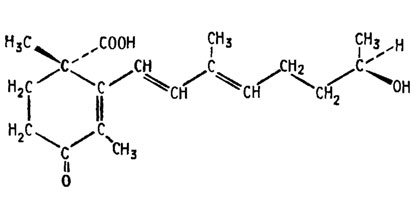
Структура триспоровой кислоты очень сходна со структурой β-ионона, а ее дипентеновая боковая цепь примерно соответствует по числу атомов углерода половине молекулы каротина, так что она является весьма вероятным прямым предшественником β-каротина. Достигаемая ее введением в среду стимуляция каротиногенеза выражается увеличением выхода продукта в 10-20 раз, например, этот выход повышается с уровня 202 мкг на 1 г массы сухого мицелия до 5200 мкг/г.
По многим данным, синтез окрашенных каротиноидов стимулируется светом. В частности, это известно для продуцирующего β-каротин Penicillium sclerotiorum. При этом в основном эффективны лучи с длиной волн от 400 до 500 нм, являющиеся областью поглощения β-каротина. Бесцветные каротиноиды, видимо, образуются без участия света (Колот, 1969). Этот спектр действия оказался также сходным со спектром фототропизма спорангиеносцев Phycomyces и колеоптилей овса, что, возможно, говорит о взаимосвязи механизма этих фототропических реакций и биосинтеза каротина (Rau, 1967). Непрерывное освещение мощностью в 1076 люкс увеличивало синтез каротиноидов у гриба Syzygites megalocarpus, который образовывал β- и γ-каротины и ликопин (Wenger, Lilly, 1966).
β-Каротин получали до последнего времени для медицинских целей из моркови. Ввиду сезонности, относительной дороговизны этого сырья для животноводческих целей и большой потребности в них в этой отрасли хозяйства проводятся многочисленные изыскания с целью получения каротиноидов микробиологическим методом. Для этого пытаются использовать розовые дрожжи из рода Rhodotorula, например Rhodotorula gracilis (Вечер, Куликова, 1968). Однако этот вид образует много каротиноидов, не могущих служить источником витамина А, как например торулородин, торулин и другие, но может одновременно служить кормовым белком. Последовательность накопления отдельных каротиноидов позволяет выяснить путь их биосинтеза, который, очевидно, достигает максимума после наибольшего накопления биомассы и складывается из последовательности: β-каротин → торулин → торулородин. Неясной остается только роль фитоина, образующегося в количестве, в 10-20 раз большем, чем другие каротиноиды.
Кроме этих организмов был обнаружен еще один перспективный продуцент каротиноидов - базидиомицет Dacrymyces deliquescens из группы фрагмобазидиомицетов. Этот гриб оказался хорошо растущим на солодовом и картофельнодекстрозном агаре, а также и на древесине.
Были сделаны также попытки использовать для биосинтеза каротиноидов Penicillium sclerotiorum, Choanephora persicaria, С. cucurbitarum и виды Neurospora.
О значении каротиноидов для грибов существует очень много разнообразных мнений, что, вероятно, связано с реальным разнообразием их функций. Иногда считают, что они принимают участие в окислительно-восстановительных процессах в клетке и поэтому связаны с митохондриями. Из этого вытекает другая приписываемая им роль экранов, защищающих клетку от избыточного окисления. При культивировании грибных организмов в условиях усиленной аэрации или на средах с дефицитом азота, где окисление идет интенсивно, они синтезируются в большом количестве.
Им также приписывалась роль половых гормонов, так как известно, что их синтез нередко связан с гаметообразованием и что у гамет одного пола они накапливаются более интенсивно, чем У другого. У мукоровых грибов их больше у женского (+) мицелия, а в гаметах Allomyces javanicus, наоборот, в подвижных мужских гаметах. Кроме этих функций они участвуют у грибов в фототропических изгибах спорангиеносцев у мукоровых, сумок у аскомицетов и плодовых тел ряда высших грибов. Мевус приписывал роль определяющего пол гормона бесцветному каротиноиду кроцетину, синтезируемому хламидомонадами. Для грибов таких сведений пока не имеется.
Хиноны, производные терпенового обмена. Из других веществ, близких по строению к каротиноидам, предположительно включающихся как кофакторы в состав ферментов дыхательной цепи, у грибов обнаруживаются производные убихинона или коэнзима Q (рис. 5.13). По-видимому, он является компонентом дыхательной цепи между цитохромом b и флавиновыми дегидрогеназами или компонентом какого-либо побочного пути окисления. Сейчас выяснено, что в процессе бактериального фотосинтеза он является передатчиком энергии фотонов с бактериохлорофилла на НАД-дегидрогеназу.
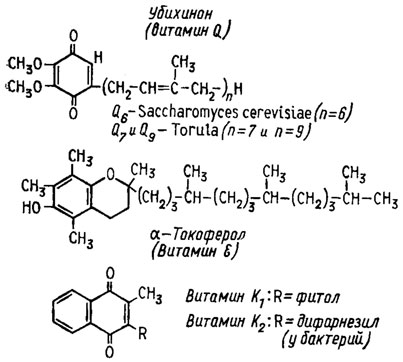
Рис. 5.13. Коферменты дыхательной цепи, образующиеся при комбинации хнноинов с изопреновыми соединениями (Девис и др., 1966)
Как и фолиевая кислота, убихиноны представляют собой группу сходных соединений, отличающихся друг от друга числом изопреновых фрагментов их длинной боковой цепи. Среди грибов эти соединения были найдены у дрожжей, в числе которых у Saccharomyces cerevisiae был найден убихинон Q6 с 6 изопреновыми фрагментами в боковой цепи, а у Torula Q7 и Q9 соответственно с 7 и 9 изопреновыми мономерами в цепи. Убихиноны с 6, 9 и 10 изопреновыми мономерами были также найдены у гифообразующих грибов, в том числе у видов родов Aspergillus, Penicillium, Neurospora, а также у Ustilago maydis и Agaricus xampestris (Lindenmayer, 1965). Относительно потребностей грибов в такого рода соединениях пока ничего неизвестно. Вероятно, они синтезируют их в необходимом им количестве самостоятельно.
Из дальнейших близких к убихинону витаминов, играющих роль в дыхательной цепи, известны токоферол (витамин Е/см. рис. 5.13) и витамин К, но относительно наличия их у грибов или потребности в них пока сведений не имеется. Витамин К2, содержащий в боковой цепи дифарнезол, был обнаружен только у бактерий (Девис и др., 1966).
Витамин D (кальциферол) и другие стероиды. Грибы, у которых особенно широко развит обмен терпенов, известны как обильный источник стероидов, занимающих важное место в их метаболизме как конструктивный элемент клеточных мембран хитинообразующих грибов, организующий у них функции проницаемости. Вероятно, они несут также ряд побочных функций.
Относительно своеобразия состава и путей биосинтеза стеролов грибов, относящихся преимущественно к группе эргостерина, уже упоминалось выше, как и о том, что основная доля их общего количества составлена провитамином D, эргостерином, из которого при облучении его ультрафиолетовым светом можно получить этот витамин в его активной форме кальциферола. Примером количественного уровня в грибах эргостерина может служить результат его анализа в мицелии Penicillium chrysogenum, полученном после ферментации пенициллина, в котором его обнаруживали в количестве до 1,1% от массы сухого мицелия. В связи с этим отходы мицелия после ферментации пенициллина одно время служили для получения витамина D и приготовления лечебного драже. В настоящее время витамин D получают для медицинских целей чаще из пекарских дрожжей. Синтез стероидов, которые у грибов, как и каротиноиды, по их локализации в клетке связаны с жирами, обычно не нуждается в предшественниках и идет весьма интенсивно.
Единственная известная у грибов потребность в стеролах обнаружена у некоторых переномицетов (Gomez-Miranda, Leal, 1965) и у растущих анаэробно дрожжей, которые нуждаются при этом в эргостерине (Fries, 1965). У ряда грибов, в частности у видов Fusarium из секции Sporotrichiella, у Stachybotrys alternans, Fusella olivacea и некоторых других найдена обильная продукция стероидов (или циклических производных фарнезола) в форме водно-растворимых сапонинов с остатками олигосахаридов, облагающих токсическим эффектом в отношении кроветворной деятельности человека и животных (алиментарно токсическая алейкия) или роста трубчатых костей (уровская болезнь/Олифсон, 1957). Олифсон приписывает им структуру циклопентанфенантренов. Вероятное экологическое значение этих веществ для грибов связано с явлениями проницаемости (Беккер, 1975).
В последнее время стали появляться сведения о важном физиологическом значении для грибов еще нескольких видов стероидов. К ним относятся β-ситостерол, стигмастерол, эргостерин и холестерин, которые оказались играющими важную роль в размножении некоторых оомицетов. К числу последних относятся виды Phytophthora и Pythium. Оказалось, что эти стероиды стимулируют у них образование зооспорангиев и органов полового плодоношения (Пыстина, 1973).
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'