2. Клеточная оболочка грибов
Довольно большую долю массы сухого мицелия грибов составляет их клеточная стенка, а именно ох 5 до 15%. Состав ее сильно варьирует и часто весьма специфичен для определенных таксономических групп грибов. Это можно видеть из примеров ее состава у представителей дрожжей, хитридиевых и мукоровых грибов - зигомицетов, приведенного в табл. 1.3 (Aronson, 1965).

Таблица 1.3. Химический состав клеточной стенки грибов в % от сухого веса их оболочки (Aronson, 1965)
* (Количество золы без фосфатов.)
Структурно оболочки грибов построены на основе двухфазной системы, в которой имеются микрофибриллы, включенные в аморфную массу матрикса. По данным электронной микроскопии, она состоит не менее чем из двух слоев с различно направленной ориентацией фибрилл. Внутренний слой обычно ориентирован по главной оси клетки, наружные - под углом к ней (рис. 1.1). У дрожжей оболочка обычно многослойная, причем в наружном толстом слое локализован маннан, во внутреннем тонком - глюкан. У водных грибов, например у Allomyces, оболочка образует ложные перегородки - псевдосепты, напоминающие спицы колеса. У сумчатых и базидиальных грибов наблюдаются настоящие перегородки - септы. У сумчатых септы между клетками обычно имеют одну простую пору, по обе стороны которой в процессе ее формирования можно видеть по паре осмофильных телец Воронина. У базидиальных грибов эти поры нередко бывают очень сложными, снабженными колпачками - парентосомами (рис. 1.2). Такие поры были найдены в плодовых телах и ризоморфах ряда высших базидиомицетов (Мооrе, 1965; Burnett, 1968). Однако до настоящего времени остается не вполне ясным, относить ли это различие в структуре септальных пор аско- и базидиомицетов к их таксономической принадлежности или к гаплоидной и гомокариотической структуре генома сумчатых и к ди- и гетерокариотической природе базидиальных грибов. Таких исследований пока еще нет, но их значение для эволюционных и таксономических построений в царстве грибов весьма принципиальное.
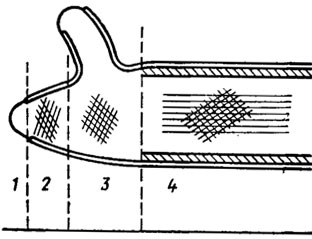
Рис. 1.1. Строение и развитие клеточной стенки грибов: 1 - ненапряженная зона кончика гифы; 2 - зона образования поперечных фибрилл методом интуссусцепции, проникновение микрофибрилл; 3 - область продольного растяжения фибрилл (область расслабления); 4 - область формирования внутреннего слоя жестких продольных фибрилл (область ригидификации и прекращения развития наружной сети фибрилл) (Burnett, 1968)
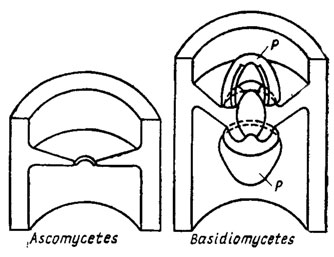
Рис. 1.2. Строение пор в клеточных перегородках вегетативных образований сумчатых и базидиальных грибов: Р - парентосомы (Мооrе, 1965)
Много нового добавили к изучению септальных пор грибов исследования на электронно-микроскопическом уровне (Flegler et al., 1976; Камалетдинова, Васильев, 1982). Во-первых, стало известно, что долипоры базидиальных грибов являются структурами, видимо, герметически изолирующими клетки мицелия друг от друга вплоть до начала образования ими плодовых тел (Flegler et al., 1976). Эта изоляция осуществляется растворяемыми протеолитическими ферментами и одновременно осмофильными (содержащими белки и липиды) двусторонними пробками, которые исчезают к моменту формирования плодовых тел. Их исчезновение сопровождается перфорациями парентосом и сообщение между клетками гиф восстанавливается.
Последние наблюдения над септами в плодовых телах дискомицетов (например, Peziza badia - Камалетдинова, Васильев, 1982) показали, что сходные разобщающие клетки гиф структуры существуют и у класса сумчатых грибов. В их формировании принимают участие тельца Воронина, образующиеся в колбообразных инвагинациях клеточной мембраны, осмофильное содержимое которых (тельце Воронина) высвобождается близ септы и располагается у отверстия септы, постепенно проникая в нее, создавая при этом замыкающее ее устройство. Дальнейшему проникновению тельца Воронина в вышележащий гимениальный слой, видимо, препятствует специальная перфорированная структура, лежащая над порой септы в материнской клетке будущей сумки, которая оказывается полностью изолированной от проникновения субмикроскопических органелл из субгимениального слоя. Подобные же изолирующие структуры наблюдаются и в формирующихся в плодовых телах парафизах.
Сходные структуры имеются и в спорулирующем мицелии несовершенных грибов, содержащих тельца Воронина, например у Arthrobotrys conoides. У продуцента пенициллина дейтеромицета Penicillium chrysogenum была обнаружена в септе структура, в точности соответствующая найденной Камалетдиновой и Васильевым у дискомицета Peziza badia (Kurilowich et al., 1980).
Скелетные основы оболочек грибов составлены кристаллически организованными полисахаридами: целлюлозой, хитином, хитозаном, маннаном, глюканами и другими. Все они имеют линейную структуру с β-1,4-связями исходных компонентов - мономеров гексоз, амино- и ацетаминогексоз. По результатам микрохимического тестирования (окраска рутениум красным) ранее считалось, что в составе клеточных оболочек грибов имеется пектин. Однако результаты химического анализа не подтвердили в ее составе входящего в структуру пектина мономера галактуроновой кислоты (Aronson, 1965).
Хитин и хитозан для большей части грибов очень характерны в составе их оболочек как азотсодержащие полимеры. При этом хитин грибов очень сходен с хитином насекомых и ракообразных, что было подтверждено картиной их дифракции в рентгеновских лучах. Однако азота в хитине грибов меньше, чем у животных, и среди его составных частей была обнаружена метилпентоза, называемая мицетозой. Хитин у грибов удается обнаружить микрохимически по методу Ван Висселинга, с помощью частичного щелочного деацетилирования и последующей реакцией на хитозан и по дифракции Х-лучей. Он не был найден только у оомицетов, таких, как сапролегниевые и пероноспоровые грибы. Ранее считалось, что хитин отсутствует у дрожжей, однако он содержится в клеточных перегородках - септах сахаромицетов (Кулаев, 1975).
В настоящее время установлено, что хитин можно обнаружить у Chitrydiales, Monoblepharidales, Protomycetales, Hyphochyiridiales, у всех Endomycetales, Blastocladiales, Mucorales, Entomophthorales, у всех сумчатых и базидиальных грибов и происходящих от них Fungi imperfecti (дейтеромицетов). Исключение составляют Oomycetes, у которых из полисахаридов в оболочке присутствует целлюлоза, полностью отсутствующая у представителей дрожжевых грибов.
В течение последнего времени в связи с выявлением возможности практического использования хитина грибов для синтеза полимеров появилось довольно много исследований с данными по более тонкому изучению его в сопоставлении с хитином ракообразных (табл. 1.4). Ацетата и D-глюкозамина меньше всего у Aspergillus niger, что объясняется более высоким содержанием пентоз и глюкозы в составе β-глюкана и двух α-глюканов оболочки гриба. Анализ дифрактограмм показал идентичную кристаллическую структуру хитина грибов и крабов с несколько большей упорядоченностью у последних. Кроме того, хитин грибов в противоположность пластинчатой структуре хитина крабов имел волокнистое строение (Феофилова и др., 1980).
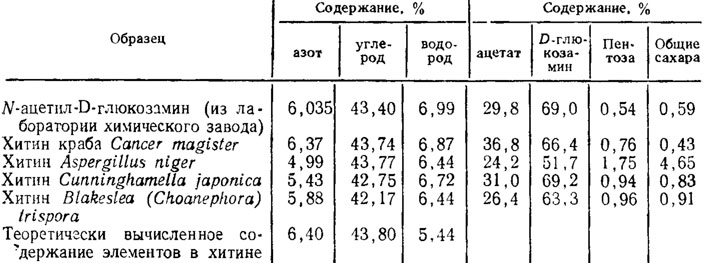
Таблица 1.4. Физико-химические свойства образцов хитина микроскопических грибов и панциря краба (% от массы образца) (Феофилова и др., 1980)
Заменяющий хитин у некоторых мукоровых грибов хитозан, обнаруживаемый при слабом кислотном гидролизе реакцией на хитозансульфат, найден, кроме Mucor rouxii, также у Phycomyces blakesleeanus. Количество ацетильных групп в нем оказалось различным и варьировало у М. rouxii до нуля. Из других аминополисахаридов полимер галактозамина со свободной аминогруппой, способной связывать фосфаты, со структурой типа хитозана, был выделен из клеточных оболочек Neurospora crassa, N. sitophitla, А. niger и Botrytls drierea (Aronson, 1965). У ряда грибов встречены полимеры из аминосахаров, ассоциированных с маннаном, глюканом и протеинами.
Целлюлоза в клеточных оболочках грибов обычно не встречается одновременно с хитином. Исключение составляет один Rhizidiomyces из порядка гифохитридиевых грибов, у которого они были обнаружены одновременно. Целлюлоза была обнаружена у грибов из оомицетов порядков Acrasiales, Lagenidiales, Saprolegniales, Leptomitales, Peronosporales (Aronson, 1965).
Среди грибов, обитающих в водной среде, целлюлоза встречается обычно только в группах, имеющих двужгутиковые зооспоры. У Blastocladiales и Monoblepharidales, имеющих одножгутиковые зооспоры, ее нет. Исключение, которое составляет Rhizidiomyces из порядка Hyphochytridiales, имеющий одновременно и хитин, и целлюлозу и считающийся по Нейбл переходной формой между имеющими хитин Chytridiales и Blastocladiales и содержащими целлюлозу оомицетами, объяснимо. Эта одножгутиковая форма имеет жгутик зооспор перистый, а не бичеподобный, как у Blastocladiales и Monoblepharidales. Любопытно, что строение ворсинок перистых жгутиков оомицетов напоминает строение жгутиков бактерий, тогда как бичеподобные полностью схожи со жгутиками флагеллат.
Целлюлоза у грибов легко обнаруживается микрохимически реактивом Шпейцера или реактивом, состоящим из раствора йода в йодистом калии с добавлением 70%-го раствора серной кислоты.
Глюканы у грибов очень обильны и отличаются от целлюлозы по их структуре. Мономеры их также представляют собой глюкозу. Однако наиболее изученные глюканы дрожжей имеют менее кристаллические структуры, чем целлюлоза. Кристалличность глюка-нов увеличивается вследствие образования гидроглюканов при их обработке серной кислотой. Подобные глюканы, находящиеся в ассоциации с протеинами, были обнаружены у дрожжей и у Penicilliunt notatum (Aronson, 1965). Другой глюкан, входящий в состав оболочек грибов, - каллоза, сходная с обнаруживаемой в ситовидных трубках высших растений и сильно красящаяся основными красителями, т. е. имеющая кислую природу, в противоположность целлюлозе имеет β-1,3-глюкозидные связи. Подобный глюкан, красящийся основными красителями, был обнаружен у Sclerofinia. Глюканы имеются также у Aspergillus fischeri, Allomyces macrogynus, Neurospora crassa. У грибов встречаются также гетерополисахариды, составленные из мономеров различных сахаров, особенно частые у рода Penicillium. У дрожжеподобных форм, патогенных для животных, как у Coccidioides и Cryptococcus, подобные же, но кислые полисахариды входят в состав окружающих их клетки капсул. Примеры глюканов и других продуктов полимеризации моносахаров и сахарных кислот, входящих в состав оболочек клеток и запасных веществ грибов, приведены в табл. 1.5.
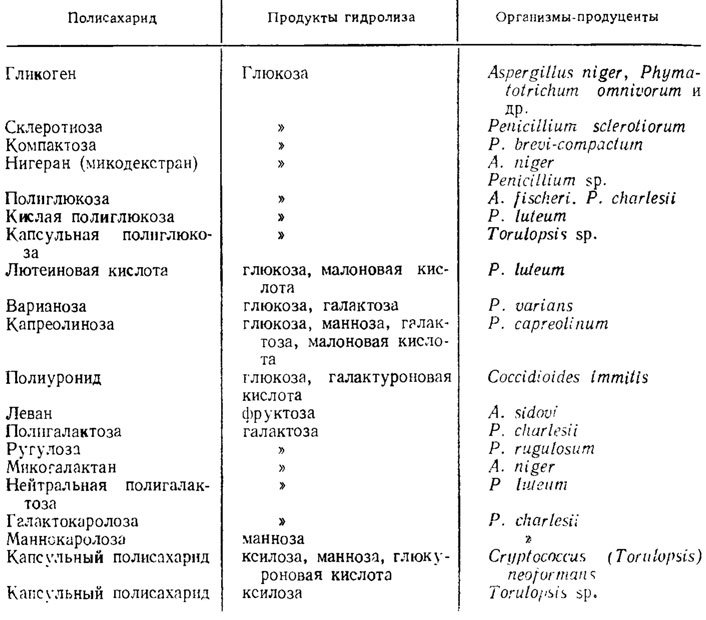
Таблица 1.5. Примеры полисахаридов грибов с расшифрованной структурой по Кокрейну (Cochrane, 1958)
Маннаны, полисахариды, составленные из мономеров маннозы. встречаются особенно обильно у дрожжей, причем они часты у видов дрожжей, обитающих на поверхности камбиального стоя под корой деревьев. К числу таких форм относится обитающая под корой хвойных деревьев Hansenula; в окружающей клетки этих дрожжей капсуле полисахарид присутствует в форме фосфоманнана. Этот гидрофильный и слизистый полисахарид прилипает вместе с дрожжами к щетинкам, покрывающим тело жуков короедов, и таким путем дрожжи переносятся с их помощью с одного дерева на другое (Wickerham, Barton, 1961). У гифообразующих грибов маннаны пока обнаружены не были, но в гидролизатaх их клеточных стенок встречается манноза.
У грибов встречаются также полисахариды, содержащие галактозу, 6-дезоксипентозу, метилпентозы, наиболее часто фруктозу, особенно у мукоровых. В оболочках клеток Penicillium chrysogenum были найдены из числа пентоз 6-дезоксипентоза, рамноза и ксилоза, которую обнаружили также у трутовика Polysiictus sanguineus.
Полисахаридно-протеиновые комплексы были обнаружены у дрожжей, например у Candida albicans - возбудителя молочницы грудных детей. Маннаново-протеиновый комплекс был найден у Saccharomyces.
Липиды у грибов количественно сильно варьируют в зависимости от условий окружающей среды и возраста культуры. Иногда их количество достигает 35-36% от массы сухого вещества клеток. У дрожжей более 3% липидов найдено в их оболочках. Обнаружены они и в оболочках мукоровых грибов, например у Мuсоr rouxii и Phycomyces, в спорангиеносцах которого установлено около 25% липидов от их сухой массы. Вероятно, они содержатся там в образованиях типа кутикулы (Aronson, 1965).
Пигменты также входят в заметных количествах в состав клеточных оболочек грибов. Пигменты, в особенности часто локализованный в виде особого слоя черный пигмент, меланин, очень часто встречаются в клеточных стенках мицелия или в оболочках спор многих грибов. Такой меланиновый слой имеется в оболочках аскоспор Neurospora tetrasperma (Aronson, 1965).
Меланин отсутствует у грибов с низкой активностью полифенолоксидазы, принимающей участие в его биосинтезе, и с преобладанием в обмене активных дегидрогеназ. Такие грибы, к которым относятся представители родов Fusarium, Trichothecium, Arthrobotris, Cephalosporium и многие другие, характеризуются чаще всего розовой или оранжевой окраской спорулирующих структур, зависящей от обилия каротиноидов, принимающих на себя роль светозащитного экрана и антиоксиданта, принадлежащую у темноокрашенных грибов меланину. Связь наличия каротиноидов с высоким уровнем активности дегидрогеназ объясняется тем, что оптимальные условия окислительно-восстановительного режима для действия дегидрогеназ совпадают с условиями, оптимальными для биосинтеза каротиноидов и других продуктов шунта терпеноидов. Каротиноидные пигменты уже не входят обычно в состав клеточных оболочек грибов, а локализуются либо в клеточной мембране, либо в каплях липидов, рассредоточенных в цитоплазме. В отдельных случаях у грибов встречаются совершенно особые пигменты, как оммохромы, глазные пигменты насекомых, у трутовика Pycnoporus (Polyporus) cinnabarinus (Шиврина, 1965) или фикобилины у видов сыроежек Russula emetica и R. paludosa (Ефименко, 1972), входящие в систему фотосинтеза у сине-зеленых и красных водорослей.
Круглосуточный доступ к анкетам на https://tumenlust.com сделает ваш поиск максимально удобным.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'