3. Основной обмен соединений углерода у грибов
Основным обменом называют этапы обмена веществ, необходимые для поддержания жизни, роста и размножения организмов. Главной чертой его является менее выраженная специфичность по сравнению с боковыми путями обмена. Он, в общем, сходен у всех живых существ и чаще отличается лишь некоторой возрастной специфичностью. Например, эмбриональные ткани получают энергию преимущественно через гликолитический путь Эмбден - Мейергофа - Парнаса (ЭМП), тогда как более зрелые характеризуются более развитым пентозным циклом или циклом гексозомонофосфата (ГМФ).
В качестве примера этого положения из области грибов могут служить прорастающие хламидоспоры Ustilago maydis, которые через 12 ч после начала их проращивания еще не имеют энзимов для усвоения Сахаров через путь ГМФ и ферментов цикла Кребса, но уже обладают комплексом энзимов, участвующих в усвоении Сахаров по пути ЭМП (Blumental, 1965). В спорах Tilletia caries имеется только серия энзимов для метаболизма Сахаров по пути Энтнер - Дудорова (ЭД), в котором отсутствует фосфорилирование глюкозы, тогда как в мицелии вместо них обнаруживаются энзимы, действующие при метаболизме Сахаров через пути ЭМП и ГМФ.
У больших таксономических групп, стоящих на сильно удаленных друг от друга ступенях филогенеза, степень развития отдельных этапов основного обмена может также довольно резко отличаться. Например, у оомицетов цикл Кребса, видимо, развит слабо, так как они используют предпочтительно для питания уже готовые аминокислоты. У паразитов часто выявляются вторичные дефекты, связанные с паразитарным образом жизни.
Хотя у грибов и встречаются исключения экологического происхождения в форме сильных изменений первых этапов основного обмена, например сильное преобладание в обмене Caldariomyces fumago редко встречающегося типа гликолиза по пути ЭД, однако основной обмен меняется реже в зависимости от экологических условий, чем его боковые пути. Он обычно не имеет видовой или групповой специфичности, характерных для узких таксономических групп грибов, обитающих в сходных экологических условиях. Видовая или групповая специфичность обмена проявляется чаще всего в закрепленных филогенетически приспособлениях, развившихся в результате экологических воздействий в боковых ветвях метаболизма, необязательных для роста и развития, накопления биомассы и деления клеток, но необходимых для приспособления к среде обитания, как например, образование антибиотиков, токсинов, витаминов, ферментов специального назначения, чаще всего гидролитических и других специфических продуктов метаболизма.
Пути гликолиза у грибов. В настоящее время исследовано четыре вида путей гликолиза у грибов, являющихся первыми этапами их углеродного обмена. Эти пути следующие: путь ЭМП, путь ГМФ или пентозный (апотомический) цикл, путь ЭД и глюкуронатксилулозный (ГК). Не исключается открытие и новых путей гликолиза у грибов.
Путь ЭМП, который был изучен первым и оказался наиболее широко распространенным, характерен промежуточным метаболизмом через образование на первом этапе фруктозо-1,6-дифосфата (рис. 3.1). На втором и параллельном ему третьем этапах происходит дальнейшая трансформация триоз, полученных после действия альдолазы.
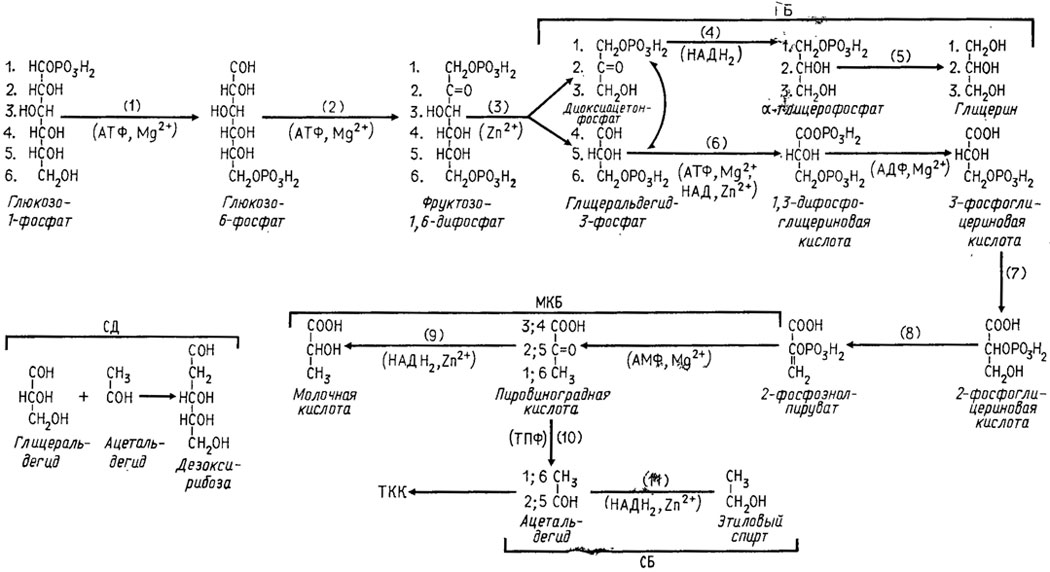
Рис. 3.1. Метаболизм глюкозы через путь ЭМП. Участвующие в цикле ЭМП энзимы: 1 - гексокиназа; 2 - фосфоглюкокиназа и глюкозофосфатизомераза; 3 - альдолза; 4 - диоксиацетонфосфатдегидрогеназа; 5 - глицерофосфатаза; 6 - глицеральдегид-3-фосфатдегидрогеназа; 7 - фосфоглицераткиназа; 8 - энолаза (фосфоэнолпируватгидратаза); 9 - козимаза; 10 - декарбоксилаза; 11 - козимаза. Кофакторы энзимов показаны в местах их действия. Распределение атомов углерода глюкозы обозначено порядковыми номерами
Пировиноградная кислота - конечный продукт процесса гликолиза - включается далее в метаболизм цикла ТКК. Однако еще не доходя до цикла ТКК, у многих грибов пировиноградная кислота может восстанавливаться при помощи НАДН2-дегидрогеназы до молочной кислоты. Это превращение носит название молочнокислого брожения.
Четвертый этап составляет превращение пировиноградной кислоты при действии декарбоксилазы с кофактором тиаминпирофосфатом (ТПФ) в ацетальдегид и далее при действии НАДН2-дегидрогеназы в этиловый спирт и носит название спиртового брожения.
Пятая трансформация в пределах пути ЭМП - это образование путем конденсации ацетальдегида и глицеральдегида сахарного компонента ДНК - дезоксирибозы, очень необходимой для основного обмена нуклеиновых кислот.
Путь гликолиза через ГМФ, или пентозный цикл, осуществляется через образование из глюкозы на первом этапе глюконолактона и из него глюконовой кислоты (рис. 3.2).
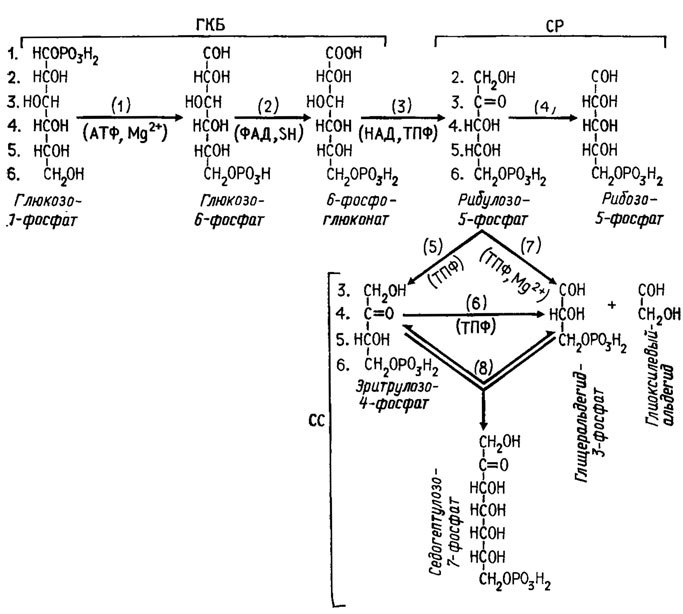
Рис. 3.2. Метаболизм глюкозы через путь ГМФ, или пентозный цикл. Участвующие в пути ГМФ энзимы: 1 - гексокииаза; 2 - глюкозофосфатоксидоредуктаза; 3 - фосфоглюкоиатдегидрогеиаза и декарбоксилаза; 4 - рибулозо-5-фосфатизомераза; 5 - рибулозо-5-фосфатдекарбоксилаза; 6 - эритрулозо-4-фосфатдекарбоксилаза; 7 - траискетолаза; 8 - траисальдолаза или траискетолаза. Распределение атомов углерода глюкозы обозначено порядковыми номерами
Второй этап пентозного цикла состоит в трансформации рибулозо-5-фосфата по двум альтернативным путям (рис. 3.2). Фермент траискетолаза, способный переносить с одного сахара на другой двухфрагментные группы углеродных соединений, содержащие кетогруппу, очень типичен для пути ГМФ.
Третий этап пентозного цикла состоит в конденсации с помощью трансальдолазы эритрулозо-4-фосфата и глицеральдегид-3-фосфата в сахар седогептулозу, функционирующую как резервный источник для образования серии Сахаров с различным числом атомов углерода. Седогептулоза может образовываться также путем переноса двууглеродного фрагмента транскетолазой на пентозу. Возможны и другие перекомбинации, осуществляемые транскетолазой и трансальдолазой.
Цикл метаболизма по пути ГМФ распространен почти так же широко, как и цикл ЭМП, и имеет весьма разнообразное назначение, поскольку: 1) он необходим для образования рибозы, а следовательно, для синтеза РНК, 2) с его помощью создаются запасы разнообразных сахаров, пригодных для дальнейших перестроек (седогептулоза) и синтезов, 3) он обеспечивает материалом биосинтезы ароматических аминокислот из эритрозо-4-фосфата.
Путь метаболизма глюкозы по Энтнер - Дудорову был обнаружен в связи с выявлением видов грибов, совершенно лишенных фосфогексокиназы, фермента фосфорилирующего гексозы. Caldariomyces fumago и прорастающие споры головни Tilletia caries оказались способными окислять нефосфорилированную глюкозу в глюконовую кислоту с помощью глюкозооксидазы и фосфорилировать уже глюконат при действии глюконаткиназы (рис. 3.3). Конечные продукты цикла ЭД могут далее включаться в циклы ЭМП и ГМФ, а также в цикл трикарбоновых кислот. Этот путь метаболизма углеводов встречается у грибов сравнительно редко.
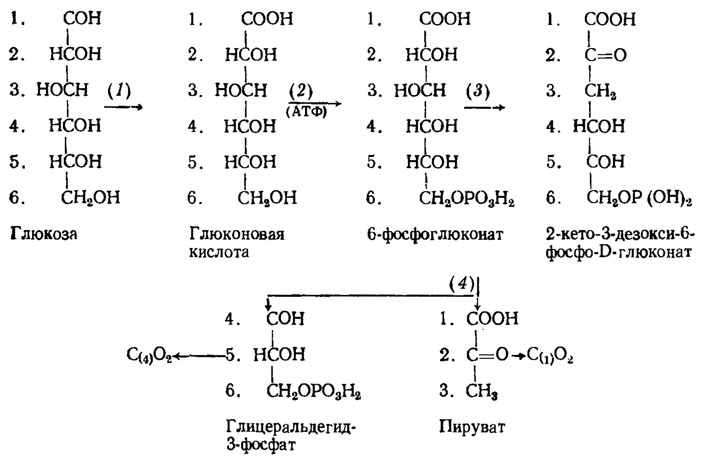
Рис. 3.3. Метаболизм глюкозы через путь ЭД и распределение в его продуктах атомов углерода этого моносахарида (обозначены порядковыми номерами). Участвующие в пути ЭД энзимы: 1 - глюкозооксидаза; 2 - глюконаткиназа; 3 - 5-фосфоглюконатдегидрогеназа; 4 - 2-кето-3-дезоксиглюконатальдолаза (Blumental, 1965)
Глюкуронат-ксилулозный путь метаболизма углеводов заключается в окислении глюкозы в глюкуроновую и далее гулоновую кислоту, дающую затем начало аскорбиновой кислоте (рис. 3.4). D-рибулозо-5-фосфат может включаться в цикл ГМФ.
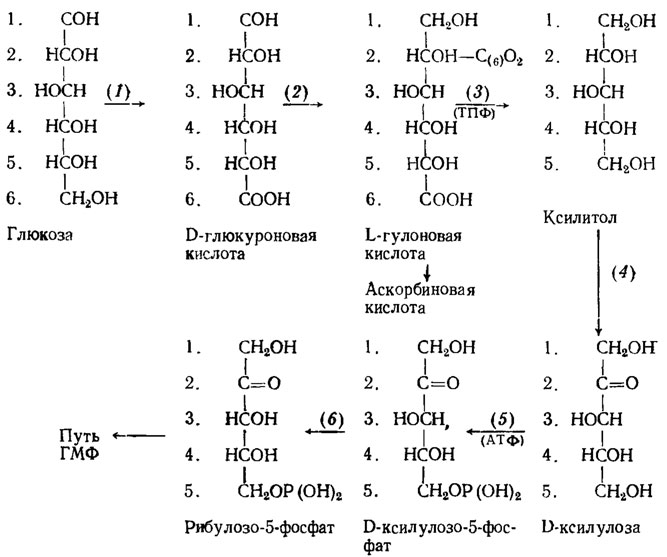
Рис. 3.4. Метаболизм глюкозы через глюкуронат-ксилулозный путь и распределение в продуктах метаболизма ее атомов углерода (обозначены порядковыми; номерами). Участвующие в пути ГК энзимы: 1 - глюкозодегидрогеназа; 2 - глюкуронатредуктаза; 3 - гулонатдекарбоксилаза; 4 - ксилитолдегидрогеназа; 5 - фосфокиназа; 6 - изомераза (Blumental, 1965)
Этот путь метаболизма имеется у Aspergillus niger, так как добавление в его культуру глюкуронолактона вызывает у него увеличение количества аскорбиновой кислоты втрое, выход которой составляет 2,6%. Рост мицелия при этом увеличивается всего на 18% (Sastry, Sarme, 1957). Количественное значение этого пути метаболизма пока еще не изучено и, вероятно, он является "минорным", поскольку объем его невелик. Тем не менее значение его в обмене важно как ведущего к образованию аскорбиновой кислоты. Кроме А. niger он, видимо, функционирует также у Penicillium chrysogenum и у дрожжей.
Описанные здесь четыре типа гликолитических процессов, вероятно, еще не исчерпывают всего разнообразия первых этапов углеродного метаболизма грибов. Например, известен в настоящее время путь окисления галактозы в галактуроновую кислоту, далее трансформирующуюся в галактодиальдозу. Такой путь окисления был обнаружен у дереворазрушающего базидиомицета Polyporus circinatus (Blumental, 1965). Вероятно, его обнаружения следует ожидать и у других видов.
Между отдельными путями гликолиза имеется связь, позволяющая обойти через включение альтернативных путей обмена затруднения из-за дефицита какого-либо энзима. Например, дефицит включающейся в обычный путь ЭМП гексокиназы у Caldariomyces fumago компенсируется возможностью исходить из образующегося без фосфорилирования глюкозы 6-фосфоглюконата или 2-кето-6-фосфоглюконата с последующим включением продуктов обмена в путь ЭМП или ГМФ (рис. 3.5).
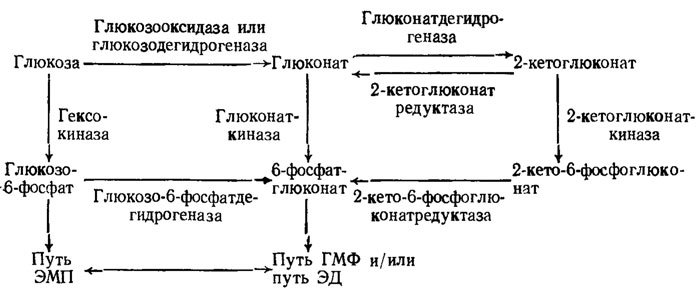
Рис. 3.5. Взаимосвязь различных путей метаболизма гексоз и гексоновых кислот у грибов (Blumental, 1965)
Количественное соотношение разных путей гликолитического обмена у различных грибов отличается друг от друга, но большей частью у них преобладает путь ЭМП. Путь ГМФ известен как ведущий, или равноценный пути ЭМП, у немногих видов, например Penicilllum charlesii, P. urticae, Ustilago maydis, Tilletia contraversa и у ряда актиномицетов.
Путь ЭД известен пока только у двух видов: С. fumago и Т. caries. Однако возможно, что впечатление о преобладании в гликолитическом обмене пути ЭМП является результатом краткосрочных опытов или других условий культивирования, так как есть сведения, что путь ГМФ развивается позднее, чем путь ЭМП, в процессе дифференцировки вегетативного мицелия.
Преобладание тех или иных путей гликолиза может очень сильно зависеть от композиции питательной среды и степени доступности необходимых для гриба энергодающих систем, минеральных компонентов, микроэлементов или витаминов. В частности, это, видимо, сильно связано с относительными концентрациями серы и фосфора в среде. Поскольку путь ЭМП основан на образовании двукратно фосфорилируемых соединений, как глюкозо-1,6-дифосфат и фруктозо-1,6-дифосфат, для него требуется более интенсивное снабжение фосфатом. Напротив, для пути ГМФ, по которому фосфорилирование протекает только однократно при образовании глюкозо-6-фосфата, фосфора требуется меньше, но зато необходимо повышенное снабжение сульфатом ввиду участия в нем SH-групп и тиамина.
Из работы Дмитриевой (1967) на примере Penicillium chrysogenum следует, что сдвиг баланса Р:S в пользу фосфора приводит к замедлению развития миделия, видимо, сопровождаемому преобладанием пути ЭМП, а сдвиг его в пользу серы - к ускорению развития с очевидным преобладанием в гликолитическом обмене пути ГМФ. Косвенными гистохимическими доказательствами наличия метаболизма по пути ЭМП при избытке фосфора в среде являются большое накопление в клетках мицелия P. chrysogenum волютина, состоящего из полифосфатов, и стимуляция синтеза ДНК, выражающаяся в учащении деления ядер. Усиление обмена ДНК объясняется синтезом по пути ЭМП дезоксирибозы, которая является одним из факторов, лимитирующих биосинтез этой нуклеиновой кислоты, а накопление полифосфатов сигнализирует о высоком уровне фосфорилирования и расхода фосфорной кислоты в обмене (рис. 3.6).
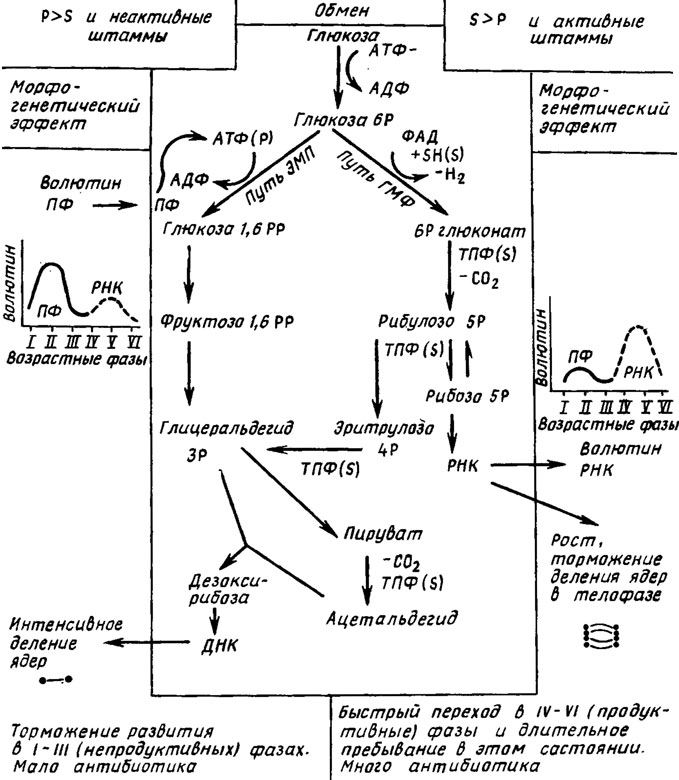
Рис. 3.6. Связь морфогенеза, гликолитического обмена и активности пеницилина в погруженной культуре Penicillium chrysogenum с преобладанием фосфора или серы в среде
Подтверждением преимущественного метаболизма по пути ГМФ при избытке серы в среде является преобладание в мицелии продуцента пенициллина волютина, состоящего из РНК, что связано с усилением синтеза рибозы, одного из важнейших факторов, регулирующих биосинтез рибонуклеиновой кислоты. Это совпадает с одновременной значительно большей продукцией серосодержащего антибиотика P. chrysogenum - трипептида пенициллина, включающего в состав молекулы цистеин. В условиях нормального оптимального баланса Р:S состояния мицелия, характерные для преобладания фосфора в среде и для сдвига баланса в пользу серы, последовательно сменяют друг друга в онтогенезе, что указывает на связь пути ГМФ (по мнению биохимиков, филогенетически более позднего) с возрастной дифференциацией и развитием экологических приспособлений в форме продукции антибиотика.
Сходные явления наблюдались и у актиномицетов, продуцентов тетрациклинов, например у Streptomyces rimosus. Низкая продуктивность сопровождалась у него угнетением цикла ГМФ, выражающимся в падении активности фосфоглюконатдегидрогеназы, и усилением пути обмена по ЭМП, о чем можно было судить по усилению активности его энзимов и по накоплению пирувата в среде.
Энергетическое значение путей гликолиза относительно невелико. Общее их назначение состоит в основном в подаче метаболитов для дальнейших биосинтезов, необходимых для нормального обмена. Вещества эти могут быть следующими: 1) составляющие фрагменты, необходимые для синтеза клеточных оболочек: глюкозамин-6-фосфат или УДФАГ для биосинтеза хитина, УДФ-глюкоза для биосинтеза глюканов, гликогена и клетчатки, манноза при синтезе маннана, сахара, участвующие в первых этапах гликолиза; 2) пентозы, необходимые для синтеза нуклеиновых кислот: дезоксирибоза для биосинтеза ДНК, образующаяся по пути ЭМП, рибоза для биосинтеза РНК, образующаяся по пути ГМФ; 3) запасные вещества для эндогенного получения энергии: УДФ-глюкоза, гликоген, жиры, глицерин и ацетальдегид; 4) метаболиты для синтезов в пределах цикла трикарбоновых кислот: ацетальдегид и пируват - источники для биосинтеза аминокислот и белка; 5) метаболиты для синтеза ароматических аминокислот: эритрулезо-4-фосфат, синтезирующийся по пути ГМФ; 6) метаболиты для биосинтеза терпенов, жиров и ароматических соединений, не содержащих азота, и т. д., такие как пируват, ацетальдегид и некоторые вещества, образующиеся по пути ГМФ.
Гликолитические пути метаболизма слабо функционируют как источники энергии, так как выход энергии в них в 20-30 раз ниже, чем в дыхательной цепи, связанной с циклом Кребса. Они играют в основном конструктивную роль в обмене веществ, поставляя разнообразные низкомолекулярные фрагменты для обеспечения различных более сложных синтезов в процессе роста и развития живых организмов, в том числе и для многочисленных боковых путей обмена, лежащих в основе их приспособлений к среде обитания.
Цикл ди- и трикарбоновых кислот Кребса. Необходимость этапа основного обмена через цикл Кребса обусловлена тем, что он дает основной материал для синтеза протеина, нуклеиновых кислот (пуринов и пиримидинов) и различных кофакторов ферментов белков, т. е. витаминов. Реакции цикла Кребса широко распространены среди грибов. Они установлены у 51 вида из всех основных классов грибов (Niederpruem, 1965), но интенсивность его функционирования различна у разных представителей этого царства. Также, видимо, не у всех форм грибов представлены все реакции цикла ТКК.
Была сделана попытка связать распространение этого обмена с филогенией грибов. При этом обнаружилось, что среди низших грибов более примитивные одножгутиковые формы характеризуются гомоферментным обменом, т. е. образуют большей частью из глюкозы молочную кислоту, и имеют слабо функционирующий цикл ТКК. Исключение составляет только род Attomyces - выраженный аэроб с сильно развитым циклом Кребса. Однако двужгутиковые оомицеты из семейств Saprolegniales и Leptomicetales тоже слабо образуют органические кислоты, формирующиеся в цикле ТКК.
Исследование обмена по циклу Кребса у грибов проводится теми же путями, что и при изучении процесса гликолиза. Оценка по анализу ферментативных систем цикла ТКК затрудняется тем, что большинство его ферментов, в противоположность ферментам путей гликолиза, находящимся в цитоплазме, локализуется в органоидах клетки - митохондриях, легко разрушающихся при их изоляции в бесклеточных экстрактах грибов. При работе с этими ферментами потребовалась разработка специальных мягких методов выделения митохондриальных фракций клеток и последующего отделения ферментов от структурных белков митохондрий, с которыми они связаны, без потери их активности. Для этого было применено быстрое механическое разрушение клеток в подходящем растворителе, заключение их в раствор сахарозы, препятствующий набуханию митохондрий, удаление вредных металлов и т. д. Дыхательные гранулы из митохондрий, активно дышащие и красящиеся rH-индикаторами, например янусом зеленым, были выделены из Allomyces macrogynus, Candida albicans, Fusarium linir Aspergillus oryzae, Myrothecium oxysporum, Neurospora crassa и ряда других грибов (Niederpruem, 1965). У A. tnacrogynus и А. oryzae были также выделены митохондрии, способные производить окислительное фосфорилирование.
Последовательность реакций цикла дикарбоновых и трикарбоновых кислот (ТКК) или цикла Кребса помещена на схеме рис. 3.7.
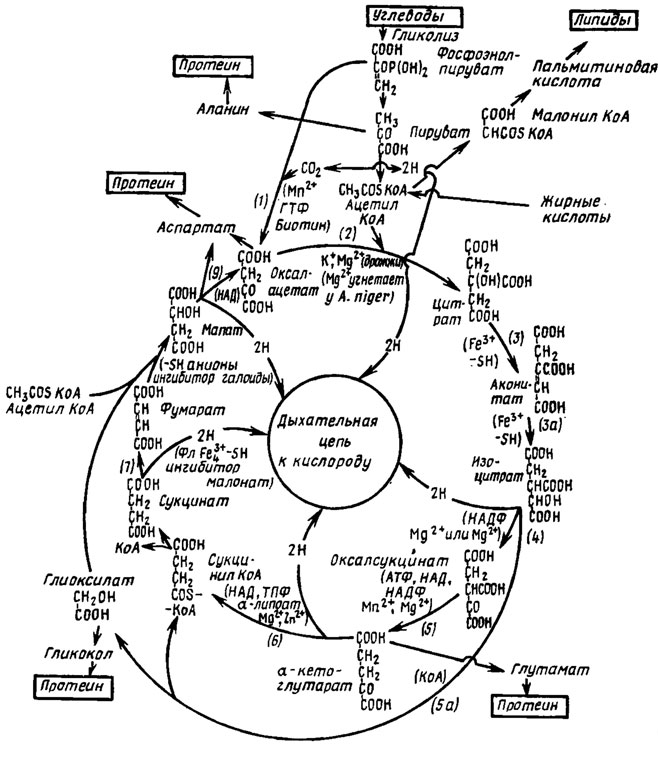
Рис. 3.7. Цикл Кребса (модификация, составлениаи по даииым Niederpruem, 1965). В скобках приведены кофакторы и стимуляторы активности данного энзима. Участвующие в цикле Кребса энзимы: 1 - пируваткарбоксилаза; 2 - цитратсиитетаза; 3 - аконитаза; 3а - аконитгидратаза; 4 - изоцитратдегидрогеназа; 5 - изоцитратдекарбоксилаза; 5а - изоцитратаза; 6 - α-кетоглутаратдегидрогеиаза; 7 - сукцинатдегидрогеназа; 8 - фумараза; 9 - малатдетидрогеназа
В первом этапе с помощью конденсирующего энзима в молекулу фосфоэнолпирувата включается молекула углекислоты, в результате чего образуется оксалацетат с четырьмя атомами углерода.
Второй этап состоит в конденсации оксалацетата с ацетильной группой с образованием шестиуглеродной лимонной кислоты. Кофакторами этой синтетазы являются коэнзим А, ионы К+ и Mg2+. Цитратсинтетаза была выделена из Aspergillus niger и из дрожжей, но у A. niger она несколько отличалась от дрожжевой, так как не стимулировалась, а угнеталась ионами Mg2+.
В третьем этапе цикла ТКК происходят внутренние перестройки, приводящие к преобразованию цитрата сначала в цисаконитовую, а затем в изолимонную кислоту. Оба этих этапа полностью обратимы. В первом из них может участвовать высокомолекулярная аконитгидратаза, не активируемая ионами Fe3+ и SH-группами. Тот же процесс трансформации цитрата в цисаконитат и обратно и последующий из цисаконитата в изоцитрат может осуществлять и другой низкомолекулярный фермент - аконитаза, активируемая ионами Fe3+ и SH-группами.
В четвертом этапе цикла Кребса изоцитрат окисляется изоцитратдегидрогеназой в оксалсукцинат. Кофакторами этого фермента являются НАДФ и металлы Мn2+ или Mg2+. При этом происходит первый сброс двух атомов водорода в дыхательную цепь с выходом энергии.
В пятом этапе цикла ТКК происходит декарбоксилирование оксалсукцината с участием связанной с НАД, НАДФ, Мn2+ или Mg2+ изоцитратдегидрогеназы, приводящее к образованию пятиуглеродной α-кетоглутаровой кислоты. Производящий эту трансформацию фермент был изучен у дрожжей и у A. niger, у которого он связан с НАД и более лабилен в отношении температуры. Ферменты этого этапа тесно связаны с митохондриями и требуют для поддержания их активности присутствия АТФ или ортофосфата. Образующийся в этом этапе α-кетоглутарат при его аминировании дает начало образованию глутаминовой кислоты, включающейся далее в синтез белков. Однако при недостатке восстановленных форм азота α-кетоглутаровая кислота метаболизируется далее.
Шестой этап трансформации метаболитов цикла ТКК состоит в декарбоксилировании α-кетоглутарата и одновременной конденсации образовавшегося сукцината с коэнзимом А. Эта реакция осуществляется с помощью α-кетоглутаратдегидрогеназы, производящей окислительное декарбоксилирование при участии в качестве кофакторов Mg2+, Zn2+, ТПФ, НАД, α-липоевой кислоты и КоА, содержащего SH-группу. При этом происходит освобождение энергии, сопровождающее второй сброс в дыхательную цепь двух атомов водорода и образование четырехуглеродного метаболита (сукцината) в цикле ТКК. Поскольку производящий эту реакцию фермент имеет в своем составе КоА и, следовательно, SH-группы, при утрате им активности под влиянием арсеиита оиа может быть возвращена при действии тиоловых соединений. Ферменты этого этапа также содержатся в митохондриях и были получены из мицелия A. niger (Ramakrishnan, 1954, цит. по Niederpruem, 1965).
В следующем, седьмом, этапе цикла ТКК сукцинил коэнзим А трансформируется в сукцииат и с помощью сукцииатдегидрогеназы, содержащей в качестве кофакторов флавииовый иуклеотид, 4 молекулы негемового Fe3+ на молекулу энзима и SH-группы, трансформируется в фумаровую кислоту. При этом происходит третий сброс двух атомов водорода в дыхательную цепь и третий случай освобождения энергии в процессе прохождения этого цикла. Сукцинатдегидрогеназа была получена в виде препарата 65% чистоты из дрожжей, а также была выделена из ряда гифообразующих грибов таких, как Aspergillus niger, Neurospora crassa, Myrothecium verrucaria, Penicillium chrysogenum и Claviceps purpurea. Энзим из С. purpurea оказался сходным с соответствующим энзимом млекопитающих по оптимальной активности при рН 7,7 и конкурентному подавлению его фумаратом, пирофосфатом и малонатом. Однако в противоположность энзиму из млекопитающих, эизим из Claviceps мало реагировал на действие фосфата.
Восьмой этап цикла Кребса состоит в гидрировании фумаровой кислоты с помощью фумаразы и в превращении ее в яблочную кислоту (малат). Фермент фумараза стимулируется двух- и трехвалентными анионами и SH-группами, но подавляется одновалентными анионами, такими как галоиды и CNS. Этот фермент был выделен из А. niger и из ряда дрожжей. У дрожжей Candida utilis он существует в двух формах с разным оптимальным рН для их активности. Обе формы удалось разделить с помощью электрофореза.
Последний, девятый, этап цикла ТКК состоит в окислении малата в оксалацетат, что осуществляется с помощью фермента малатдегидрогеиазы, которая относится к дегидрогеиазам α-гидроксикарбоксиловых кислот и имеет в качестве кофактора НАД. Эта дегидрогеназа была получена в виде сто-, двестикратно очищенного препарата из супериатанта гомогената клеток дрожжей, т. е., видимо, не связана с митохондриями. У Neurospora crassa были найдены четыре изозима этого фермента (Niederpruem, 1965), а у Uromyces phaseoli (U. appendiculatus) найдено три таких изозима в его уредоспорах. В этом этапе происходит последний, если не считать момента включения СО2 в пируват, четвертый, сброс двух атомов водорода в дыхательную цепь.
Цикл глиоксилата, являющийся укороченным вариантом цикла ТКК, обнаруженным первоначально у бактерий, встречается также и у грибов, как у дрожжевых, так и у гифообразующих. Этапы обмена по глиоксилатиому пути состоят в следующем (см. рис. 3.7).
Под влиянием фермента изоцитратазы, при одновременном участии коэизима А, происходит расщепление изолимонной кислоты на сукцинил- КоА и глиоксилевую кислоту, содержащую сочетание карбоксильной и спиртовой групп, удобное для процессов конденсации. Сукцинил-КоА трансформируется далее по ходу цикла ТКК в сукцинат и фумарат, а глиоксилат конденсируется с помощью фермента малатсинтетазы с ацетил-КоА, что приводит к образованию яблочной кислоты. При этом отпадает необходимость в прохождении трех этапов цикла ТКК от изоцитрата до сукцинил-КоА и источник для образования аспарагиновой кислоты в форме малата пополняется сразу двумя путями: через путь сукцинат-фумарат-малат и через конденсацию глиоксилата с ацетил-КоА. Кроме того, возможен третий путь через восстановление оксалацетата.
Все это приводит к быстрому накоплению обильного материала для биосинтеза аспарагиновой кислоты, получающейся путем аминирования малата или оксалацетата и вовлекающейся далее в биосинтез белков. Значение метаболизма в пределах цикла Кребса для грибов так же, как и в случае других живых организмов, сводится к трем основным функциям.
1. Синтез "материнских", или первичных, аминокислот путем аминирования кето- и оксикислот, на основе которых далее синтезируются белки. Главнейшие из этих путей аминирования состоят в трансформациях: оксалацетат (или малат)-аспарагиновая кислота, α-кетоглутарат - глутаминовая кислота и, на пороге цикла ТКК и гликолиза, пируват-аланин, а также глиоксилат-глицин.
2. Получение энергии и реализация окислительного фосфорилирования в дыхательной цепи, т. е. процесса запасания энергии. Эта функция выполняется благодаря пятикратному сбрасыванию 2Н+ в процессе окисления кислот цикла Кребса. Запас энергии накапливается при этом в форме АТФ или других макроэргических соединений и служит источником снабжения энергией всевозможных синтезов.
3. Звено цикла ТКК, в котором осуществляется усвоение СО2 при образовании щавелевоуксусной кислоты, так же, как и включение СО2 в ацетат при образовании малоната (в жировом обмене), может служить для пополнения в клетке запаса углерода, используемого в различных биосинтезах.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'