5. Процессы трансметилирования и образования аминов у грибов
В области соединений азота для грибов не менее специфичны, чем синтез мочевины, синтезы аминов и азотсодержащих спиртов, альдегидов и кислот, видимо, играющих у них экологическую роль, связанную с влиянием на проницаемость мембран. Из соединений этого типа у грибов широко распространены холин и его производные - ацетилхолин и мускарин из Amanita muscaria, бетаины (Андреева, 1967), многочисленные летучие амины базидиомицетов (List, 1958; Stein von Kamienski, 1958) и целый ряд других соединений сходного типа, как например серотонин у Paneolus campanulatus и близкие к нему по строению псилоцибин из Psylocybe mexicana (Hofmann и др., 1958, 1959) и буфотенин из видов мухомора (Wieland, Motzel, 1953).
Путь образования аминов у грибов пока не вполне ясен. Вероятно, у грибов он ближе всего к пути формирования их у животных и происходит при участии моноаминооксидазы (МАО, по терминологии медиков). Примером такого процесса может служить образование гистамина из гистидина. Процессы трансметилирования, участвующие в образовании ряда веществ типа производных холина и серотонина, служат одновременно для образования многих антибиотиков, а также из урацила или цитозина, тимина и пиримидинового компонента витамина В1. Процессы трансметилирования и биосинтез аминов связаны в едином метаболическом цикле, где донорами метильных групп являются либо пул фолиевой кислоты, либо метионин, а переносчиками метильной группы - s-аденозилметионин (SAM) или производное холина - бетаин. Детали всего обменного пула формирования аминов, азотсодержащих спиртов и метильных производных объединены в схеме на рис. 4.7.
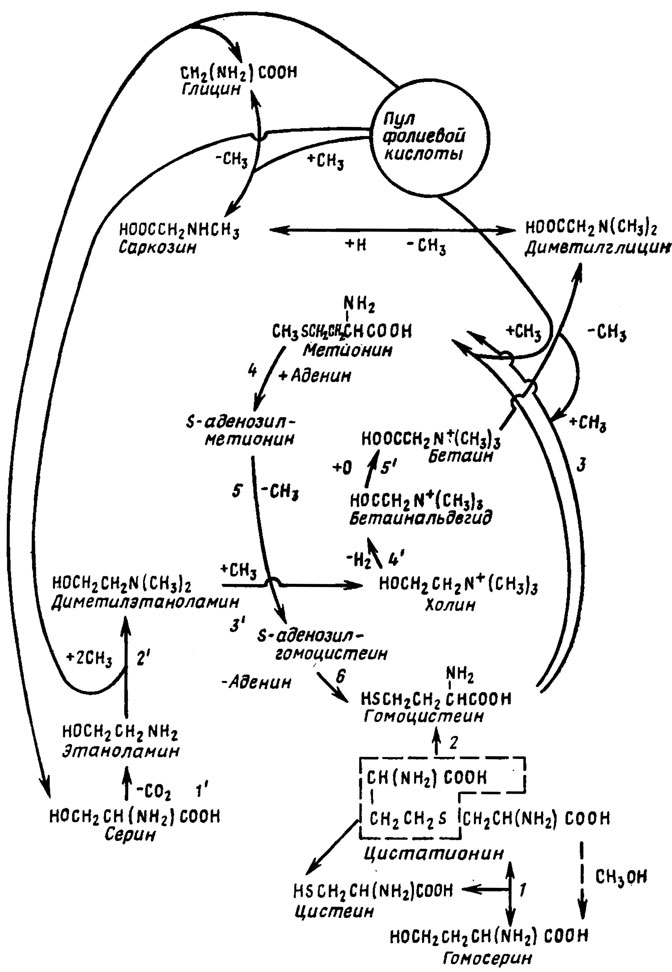
Рис. 4.7. Биосинтез аминов и холина при участии системы трансметилирования
Как показали миланские биохимики (Ciusa, Barbiroli, 1962), SAM является в организме животных агентом детоксикации очень многих токсических циклических соединений. Он функционирует и у бобовых растений, в результате чего в них накапливается тригонеллин и гомарин. Поскольку последние соединения встречаются и у грибов, видимо, реакции с участием SAM функционируют и у них.
Участие процесса трансметилирования в биосинтезе нуклеиновых кислот было обнаружено в опытах на мутантах Neurospora crassa, у которых при метиониновом голодании наблюдались снижение на 30% биосинтеза РНК и пониженный синтез белка. В качестве одного из продуктов обмена этого цикла в других опытах да мутантах N. crassa был обнаружен ацетилгомосерин, вероятно, являющийся побочным продуктом биосинтеза метионина. С биосинтезом аминокислот и простейших пептидов связано образование многих полипептидных антибиотиков и токсинов грибов, например трипептида пенициллина, дипептида лакомаразмина и других.
Как можно видеть из рис. 4.7, биосинтез аминов у грибов тесно связан с формированием бетаинов и холина, которые встречаются у них в значительных количествах. Примером этому могут служить анализы содержания бетаинов (Андреева, 1967) и холина (Корякина, 1967) в плодовых телах высших базидиомицетов. При исследовании около 60 видов этих грибов, входящих в порядок Agaricales, особенно богатыми бетаинами оказались содержащие их от 4 до 5% от сухой массы четыре вида рода Boletus, в особенности белый гриб Boletus edulis, а также два вида сыроежек и мухомор Amanita vaginata.
Холина в высших базидиомицетах оказалось примерно в 10 раз меньше, чем бетаинов, и его количество колебалось сильнее (от 0,075 до 0,9% от сухой массы). Наиболее обильно он был представлен у рода Ramaria из порядка Aphyllophorales (0,6-0,9%), у Boletus edulis и некоторых видов Lactarius (0,7-0,8%) из порядка Agaricales (Корякина, 1967).
На азотном обмене грибов довольно хорошо прослеживается вероятная эволюция их обмена в направлении усложнения участвующих в нем ферментных систем. С этих позиций их можно сгруппировать в три основные категории.
1. Формы с отсутствием системы нитратредуктазы, с редуцированным циклом Кребса и отсутствием ферментов переаминирования и глютаматдегидрогеназы. Они могут использовать только готовые наборы аминокислот. Сюда относятся в основном низшие грибы, а именно ряд форм из оомицетов и хитридиомицетов, с трудом выращиваемые в культуре.
2. Формы, способные синтезировать аминокислоты из аммиака и кетокислот, обладающие хорошо развитым циклом трикарбоновых кислот, но лишенные нитратредуктаз и отсюда слабо растущие или не растущие на средах с нитратом. Сюда можно отнести зигомицеты и некоторые низшие сумчатые грибы, например дрожжи.
3. Формы с наиболее сложным азотным обменом, имеющие вполне развитую систему нитратредуктаз и способные использовать все виды источников азота - органические, аммонийные соли и нитраты. Сюда относится большинство грибов: почти все базидиальные, высшие сумчатые и происшедшие от них дейтеромицеты.
Исключение из такого порядка иерархии составляют в основном формы со вторично упрощенным обменом вследствие обитания в условиях симбиоза или паразитизма. Системы протеолитических и запасающих энергию в форме фосфорных связей энзимов существуют у всех и начинают функционировать на наиболее ранних фазах их развития, что указывает на их наибольшую древность в филогении обмена веществ грибов.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'