3. Усвоение органического азота, синтез и использование аминокислот
Органический азот усваивается главным образом в форме аминного азота аминокислот, пептидов и белков. Белки и пептиды перед усвоением расщепляются протеолитическими ферментами, аминокислоты могут усваиваться целиком. Рост грибов лучше обеспечивается смесью аминокислот, чем отдельными из их числа. Неспособны использовать ион NH4+, и нуждаются в готовых аминокислотах некоторые хитридиомицеты и оомицеты, например Blastocladiella emersonii, Sapromyces elongatus и Leptomitus lacteus, а также Saprolegnia parasitica. Из других групп грибов нуждаются в готовых аминокислотах в основном паразиты, как Ophiobolus graminis или Trichophyton mentagrophytes. Лучше всего они используют содержащий смесь аминокислот гидролизат казеина. Дрожжи также лучше используют аминокислоты, чем ион аммония или нитрат. Бывают случаи, когда усвоение одних аминокислот угнетается присутствием в среде других (Nicholas, 1965). Усвоение аминокислот зависит также от возраста культуры, от рН культуральной среды и от концентрации в ней аминокислот, как это было показано методом изотопов для Botrytis fabae.
Аминокислоты с более короткой цепью лучше усваиваются грибами, чем с длинной. Например, аспарагин хорошо усваивается видами рода Aspergillus (Agnihotri, 1966), аланин Fusarium oxysporum (Said, Harhash, 1966). L-изомеры обычно усваиваются значительно лучше, чем D-изомеры или рацематы, так, это наблюдается у миксомицетов (Cotter, Raper, 1966) или у Phytophthora parasitica и Verticillium albo-atrum, плохо усваивающих D-аланин. D-аминокислоты не могут быть прямо включены в обмен как природные L-формы и обычно используются после их дезаминирования с помощью оксидазы D-аминокислот и трансформируются в кетокислоты, что осуществляется с различной скоростью для разных компонентов их смеси. При усвоении L-аминокислот, видимо, принимает участие система окислительного фосфорилирования, так как ингибиторы этой системы угнетают их потребление.
Такие первичные аминокислоты, как аланин и глютаминовая: кислота, могут накапливаться в мицелии Neurospora crassa в больших количествах (Aurich, 1966), так же как и в уредоспорах. Puccinia graminis, что может зависеть от отставания процесса их включения в белки по сравнению с их ассимиляцией (Nicholas, 1965). Как единственный источник азота для роста Fusarium oxysporum лучшими аминокислотами являются аспарагиновая, глютаминовая и β-аланин, т. е. первичные (Wolf, 1955). Способность использовать их как единственный источник азота может варьировать даже в пределах близких родов. Например, дрожжи из рода Saccharomyces используют как источник азота меньший круг аминокислот, чем дрожжи рода Torula. Амиды соответствующих аминокислот обычно используются лучше, чем они сами. Так, аспарагин является лучшим источником азота, чем аспартат, в частности для Tricholoma imbricatum, Piricularia oryzae, Leptographium sp. и Phycomyces blakesleeanus. Глютамин хорошо утилизируют Tricholoma gambosum и другие базидиомицеты. Аспарагин способствует развитию перитециев у Diaporthe phaseolarum. Нуждаются в органическом азоте для споруляции и некоторые другие виды, например Alternaria poonensis.
Серосодержащие аминокислоты обычно плохие источники азота, даже для нуждающихся в органическом источнике серы в виде метионина видов Allomyces и Phytophthora (Machlis, Craseman, 1956). Худшими, чем аминокислоты, источниками азота являются нуклеиновые кислоты и их производные. Из числа нуклеиновых оснований Fusarium oxysporum лучше используют пурины, чем пиримидины, пиримидиннуклеозиды, чем пиримидиннуклеотиды, и свободные основания. Так, Fusarium oxysporum лучше утилизирует цитидин и уридин, чем цитидиловую и уридиловую кислоты, и РНК лучше, чем ДНК, a Neurospora одинаково (Wolf, 1955).
Специфичная потребность в определенных аминокислотах встречается у грибов сравнительно редко. Они могут усваивать азот даже из органических нитросоединений, например из нитрированных спиртов, CN- и CNS-групп. Последние группы используют иногда паразитические грибы, если они включаются в состав алкалоидов, свойственных их растениям-хозяевам.
Еще большее влияние, чем на усвоение иона NH4+, на усвоение аминокислот оказывают дикарбоновые кислоты. Например, у Phycomyces blakesleeanus на среде с аргинином вес достигал 43 мг, а при добавке в эту среду 0,1% янтарной кислоты - 192 мг. Видимо, при этом легче происходит биосинтез набора аминокислот, образующихся в результате реакции переаминирования и участвующих далее в биосинтезе протеинов. Вторая функция дикарбоновых кислот состоит в нейтрализации получающегося при дезаминировании аминокислот избытка аммиака. Сходная причина, т. е. степень легкости использования в реакциях переаминирования, лежит в основе наблюдавшегося. Стейнбергом явления градации степени пригодности аминокислот как единственного источника азота (Steinberg, 1942). Из 22 испытанных им аминокислот наилучшими оказались 7: аланин, аспарагиновая кислота, аргинин, глицин, глютаминовая кислота, пролин и оксипролин, которые были названы первичными. Стейнберг предполагал, что остальные 15 аминокислот, входящие в состав белков, образуются на основе этих семи, что, в общем, соответствует современным представлениям.
Абсолютная специфичная потребность в отдельных аминокислотах у грибов встречается только среди мутантов, а у природных форм главным образом относительная, выражающаяся в улучшении роста в их присутствии. Например, Мuсеnа rubro-marginata реагирует так на ароматические аминокислоты, тирозин или фенилаланин, Cenococcus graniforme - на гистидин, Eremothecium ashbii - на несколько аминокислот при его росте на среде с кислым рН (Nicholas, 1965). Использование аминокислот грибами может осуществляться путем их прямого включения в белки или путем их дезаминирования и переаминирования (Фостер, 1950; Лилли, Барнетт, 1953/рис. 4.2).
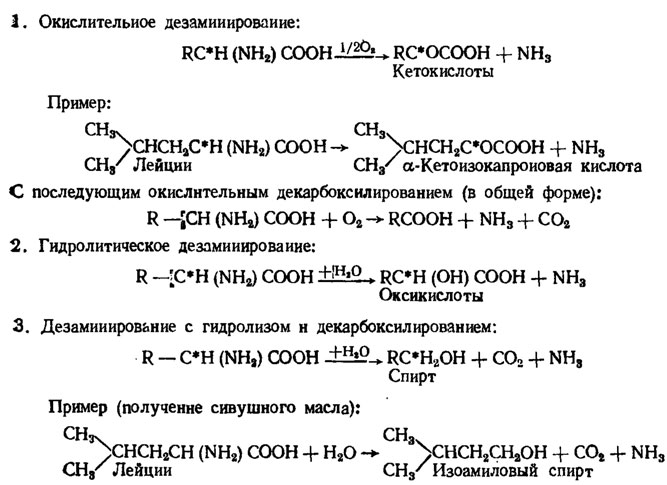
Рис. 4.2. Пути использования аминокислот грибами
Для синтеза белков цитоплазмы очень важен процесс трансаминирования (или переаминирования), который известен у многих грибов и требует в качестве кофактора пиридоксальфосфата (рис. 4.3).
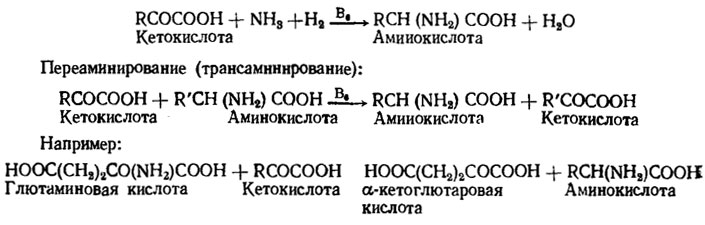
Рис. 4.3. Схема процессов синтеза аминокислот у грибов и переаминирования
Глютаминовая кислота, так же как и образующаяся из нее при декарбоксилировании γ-аминомасляная, является лучшим донором аминогрупп при переаминировании. Переаминирование с использованием этих субстратов обнаружено у очень многих видов грибов, включающих виды родов Aspergillus, Neurospora и Endomycopsis (Nicholas, 1965). Большой ряд последовательных синтезов аминокислот был изучен на моделях мутантов Neurospora к других грибов.
В последнее время аминокислотный состав плодовых тел и культивируемого мицелия грибов привлек большое внимание в связи с использованием их как пищевых и кормовых ресурсов. При этом выяснилось, что и те и другие обладают вполне полноценным; набором аминокислот, в том числе и незаменимых. Примером этого может служить работа Р. А. Масловой (1978) по анализу аминокислотного состава плодовых тел и культивируемого мицелия ряда грибов-трутовиков, среди которых ею были изучены Тrаmetes suaveolens, Cariolus zonatus, Laetiporus sulphureus, Gloeophyllum sepiarum, Lenzites betulina и Coltrticia perennis (табл. 4.5).
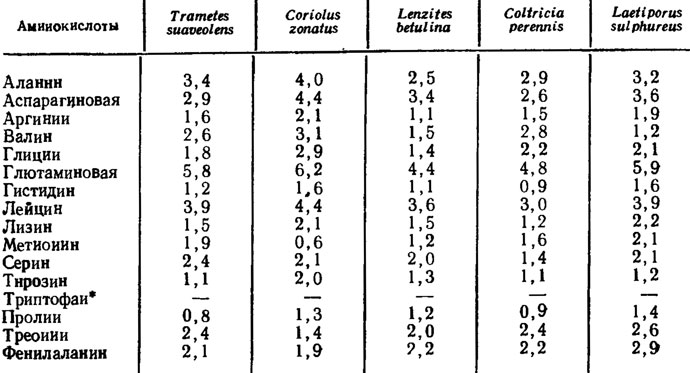
Таблица 4.5. Аминокислотный состав мицелия грибов-трутовиков, выращенных на среде с пептоном и глюкозой (в % на сухую массу) (Маслова, 1978)
* (Триптофан найден только в плодовых телах.)
При этом, что особенно важно, у культивируемого мицелия максимум накопления большинства аминокислот, судя по примеру Т. suaveolens, достигался уже на восьмые - девятые сутки роста, что практически вполне приемлемо, хотя и уступает скорости роста специально отобранного для пищевых целей штамма Polyporus squamosus А. К. Торева (1975).
Количество отдельных аминокислот в мицелии среди культур, испытанных Р. А. Масловой, достигало у преобладающих в гидролизате, как аланин, аспарагиновая и глютаминовая кислоты и лейцин, 4-5% от сухой массы мицелия. Лизина накапливалось порядка 2%. Не хватало в гидролизате культивируемого мицелия только присутствовавшего в гидролизате плодовых тел триптофана (табл. 4.5).
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://pharmacologylib.ru/ 'Библиотека по фармакологии'